











Infectious Disorders
Arboviral encephalitis
May. 15, 2026
MedLink, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Worddefinition
At vero eos et accusamus et iusto odio dignissimos ducimus qui blanditiis praesentium voluptatum deleniti atque corrupti quos dolores et quas.
The Japanese encephalitis virus is a flavivirus and a mosquito-borne human pathogen. It is the world’s most important cause of viral encephalitis and is present in many countries in Asia, causing nearly 68,000 clinical cases every year. The case fatality rate can be as high as 30%. Permanent neurologic or psychiatric sequelae can occur in 30% to 50% of patients. Children are more frequently and severely affected. A paradigm shift in epidemiology has been observed as Japanese encephalitis is becoming increasingly common among adults due to expanding Japanese encephalitis transmission areas, improved control in children leading to more adult cases, and increased tourism to Japanese encephalitis risk areas. The clinical features of Japanese encephalitis virus infection range from a nonspecific flu-like illness to a severe and often fatal meningoencephalomyelitis. MRI in patients with neuroinvasive disease characteristically shows high-signal lesions in the thalamus and substantia nigra. A report from China recorded 47 patients with Japanese encephalitis who presented with Guillain-Barré syndrome. In the majority, Guillain-Barré syndrome was of the acute motor axonal neuropathy variety. A study of 1626 patients with Japanese encephalitis (2016–2020) found 230 (14%) with peripheral nerve injury, leading to higher mortality (16%) and longer hospital stays (105). Japanese encephalitis infection can also trigger anti-N-methyl-d-aspartate receptor (NMDAR) immunoglobulin G (IgG) synthesis. Japanese encephalitis may rarely present with symmetrical quadriplegia or transverse myelitis, requiring high suspicion in atypical cases and travelers returning from endemic areas. Early diagnosis and supportive treatment aid recovery. No specific antiviral therapy is available for Japanese encephalitis. In a small trial, combination immunotherapy that included immunoglobulin, ribavirin, and interferon alpha 2b was effective. Treatment is otherwise mainly supportive and symptomatic. Intravenous immunoglobulin has been shown to augment the development of neutralizing antibodies in patients with Japanese encephalitis, and it may warrant use as a therapeutic agent in the future. Vaccination of the population at risk is the method of choice for prevention. The inactivated mouse brain-derived vaccines have now been replaced by cell culture–based vaccines. In this article, the author has reviewed in detail the various aspects of Japanese encephalitis.
|
• Japanese encephalitis virus is a neurotropic flavivirus that is the causative agent of the major mosquito-borne encephalitis in the world. | |
|
• Among 68,000 annual cases of Japanese encephalitis, approximately 30% die, and up to 50% of survivors may have sequelae. | |
|
• Japanese encephalitis, in the endemic areas, is regarded as a disease of children. | |
|
• Japanese encephalitis dominantly affects the thalamus, corpus striatum, brainstem, and spinal cord. | |
|
• Japanese encephalitis manifests with altered sensorium, seizures, and focal neurologic deficit. | |
|
• Japanese encephalitis should be considered as a diagnostic possibility in travelers developing encephalitis after travel to endemic areas. | |
|
• In the absence of effective antiviral therapy, Japanese encephalitis treatment is symptomatic and supportive. | |
|
• The main measure for Japanese encephalitis prevention is the use of a live attenuated vaccine for humans. |
Since the end of the 19th century, Japanese encephalitis has been recognized as a scourge of the orient. Epidemics of encephalitis have been described in Japanese literature since 1870, with thousands of cases recorded in some years. In 1933 a filterable agent was transferred from the brain of a fatal case and used to cause encephalitis in monkeys; the virus was then isolated in 1935. The term “Japanese B encephalitis” was used originally to distinguish these summer epidemics from Von Economo's “encephalitis lethargica” (labeled type A) (45). The term “B” has since been dropped. The virus was subsequently classed as a member of the genus flavivirus (family Flaviviridae), named after the prototype Yellow fever virus (in Latin, flavus means yellow). Although of no taxonomic significance, the ecological term “arbovirus” is often used to describe the fact that Japanese encephalitis virus is arthropod (insect) borne. A vaccine developed by Albert Sabin (later of poliomyelitis fame) and others during World War II has been available for 30 years. Despite the existence of this vaccine, Japanese encephalitis has grown as a problem in the last 50 years because of its geographical spread and increased incidence. The disease threatens both residents and travelers in endemic areas and is one of the most important emerging arboviruses (89).
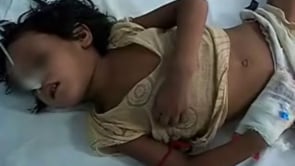
The clinical features of Japanese encephalitis virus infection range from a nonspecific flu-like illness to a severe and often fatal meningoencephalomyelitis. Children with Japanese encephalitis typically present after a few days of febrile illness, which may include coryza, diarrhea, and rigors. This is followed by headache, vomiting, and a reduced level of consciousness often heralded by a convulsion.
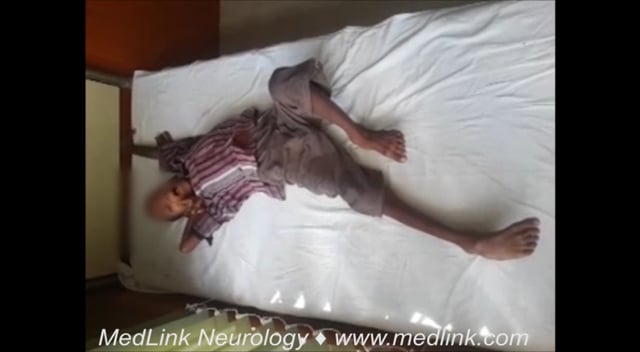
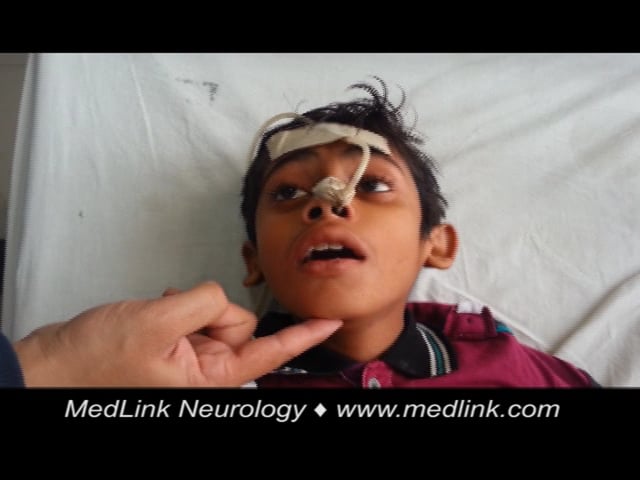
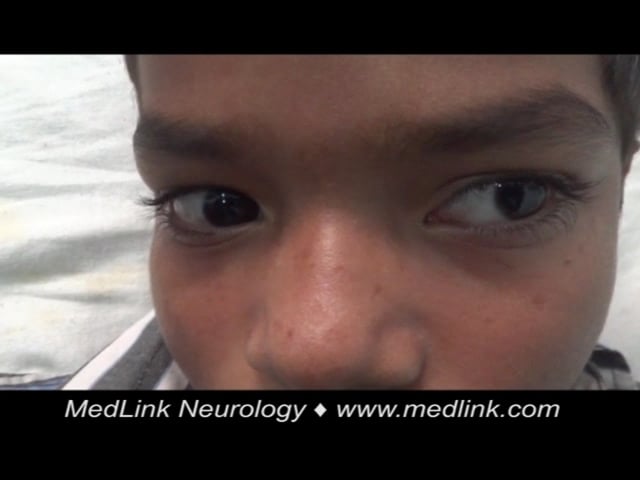
In a proportion of patients, recovery is rapid and spontaneous (“abortive encephalitis”). Others present with aseptic meningitis and have no encephalopathic features. The classical description of Japanese encephalitis includes a dull, flat “mask-like” face with wide unblinking eyes, tremor, generalized hypertonia, and cogwheel rigidity. These parkinsonian features were reported in 20% to 40% of Asian children and 70% to 80% of American service personnel (61; 54; 24). Rigidity and spasms, particularly on stimulation, occur in about 15% of patients and are associated with a poor prognosis (54). Other extrapyramidal features include head nodding and pill rolling movements, facial grimacing, lip-smacking, opisthotonus, and choreoathetosis (54; 90).
Convulsions are common in Japanese encephalitis, occurring in up to 85% of children and 10% of adults (25; 54). Multiple or prolonged seizures, including subtle motor status epilepticus, are linked to poor outcomes (96; 91). A comparative study showed that neck stiffness, convulsions, abnormal behavior, seizures, and elevated aspartate transaminase were more frequent in children, whereas cerebrospinal fluid abnormalities were more common in adults (09). In a large adult cohort, key features included altered sensorium, seizures, headache, movement disorders, brainstem signs, and severe systemic complications (82).
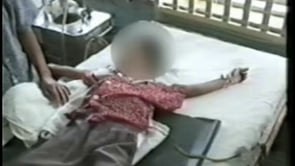
A subgroup of patients with Japanese encephalitis presented with a polio-like acute flaccid paralysis presentation (94). In these patients, a short febrile illness was followed by a rapid onset of flaccid paralysis in one or more limbs despite a normal level of consciousness (23). Weakness occurred more frequently in the legs than the arms and was usually asymmetrical. Thirty percent of such patients subsequently developed encephalitis, with reduced level of consciousness and upper motor neuron signs, but in the majority, acute flaccid paralysis was the only feature. At follow-up there was persistent weakness and marked wasting in the affected limbs. Nerve conduction studies demonstrated markedly reduced motor amplitudes, and electromyography showed chronic partial denervation, suggesting anterior horn cell damage (94). Flaccid paralysis also occurs in patients with “classical” Japanese encephalitis. It is reported in 5% to 20% of comatose patients (25) and is associated with abnormal signal intensity on T2 weighted MRI images of the spinal cord (57).
In addition to its ability to infect anterior horn neurons, Japanese encephalitis virus can be associated with cases of Guillain-Barré syndrome (104; 105). Wang and colleagues reported data on 47 patients with Japanese encephalitis who presented with Guillain–Barré syndrome (104). CSF evaluation, in 38 patients, revealed albumin-cytologic dissociation. In the majority (47%) of patients, Guillain-Barré syndrome was of acute motor axonal neuropathy variety. In 18 cases Guillain-Barré syndrome was of acute motor-sensory axonal neuropathy type. Acute inflammatory demyelinating polyneuropathy and acute sensory neuropathy verities were noted in four and three patients, respectively. Twenty-eight patients were treated with intravenous immune globulin. Half of these responded well to intravenous immune globulin treatment (104).
A report from Indonesia revealed that Japanese encephalitis virus infection was even prevalent in patients with non-encephalitic acute febrile illness (66). Serum specimens of 144 patients with acute febrile illness were tested for anti-Japanese encephalitis virus IgM antibody. Twenty-six (18%) patients either had confirmed anti-Japanese encephalitis virus IgM antibody positivity or demonstrated equivocal results. Five (3.5%) and eight (5.6%) patients, respectively, were able to fulfill the criteria for confirmed or probable Japanese encephalitis infection (66).
Japanese encephalitis can rarely present with spinal cord syndromes, such as symmetrical quadriplegia or transverse myelitis, complicating diagnosis, especially in nonendemic travelers and atypical neurologic presentations. Basnet and colleagues reported a 30-year-old man from Nepal with fever, altered mental status, and symmetrical flaccid quadriplegia (06). Cerebrospinal fluid analysis showed elevated protein and pleocytosis, and imaging revealed left temporal lobe hypodensity. Although initial herpes testing was negative, Japanese encephalitis was confirmed via positive IgM serology. The patient was treated supportively and showed gradual neurologic recovery. In a second case, Hahn and colleagues described a 37-year-old unvaccinated American who developed back pain, lower limb weakness, and urinary retention 3 days after returning from a surfing trip to Indonesia (32). Spinal MRI revealed cord edema from T10 to L1. A broad infectious workup was negative except for Japanese encephalitis, confirmed by serum and CSF IgM positivity and rising neutralizing antibodies. Treatment with corticosteroids and plasmapheresis led to partial functional recovery.
The case-fatality rate among those with symptomatic Japanese encephalitis can be as high as 30%. Permanent neurologic or psychiatric sequelae can occur in 30% to 50% of patients (12; 110). Fixed flexion deformities of the arms, and hyperextension of the legs with “equine feet” are common. Twenty percent of patients have severe cognitive and language impairment (most with motor impairment too) and 20% have further convulsions (43). A higher rate of sequelae is reported for children than adults (85). More detailed studies have suggested that approximately half of those who were classed in the “good recovery” group have more subtle sequelae such as learning difficulties, behavioral problems, and subtle neurologic signs (55). The long-term prognosis of Japanese encephalitis was assessed in Malaysia. In this study 118 patients were recruited. Ten patients (8%) died during the acute phase of illness. At hospital discharge, 44 (41%) of the 108 patients who survived had apparent full recovery; 33 (31%) patients had severe neurologic sequelae (76). On long-term follow-up, more than one half of the patients continued to experience neuropsychological sequelae and behavioral disorders. A combination of poor peripheral perfusion, with prolonged capillary refill time (greater than 2 seconds), Glasgow coma score (8 or less), and two or more witnessed seizures predicted a poor long-term outcome with 65% sensitivity and 92% specificity (76).
Chen and colleagues evaluated long-term neurologic complications and mortality in 352 adult patients with Japanese encephalitis (17). Stroke was the most common neurologic complication, followed by epilepsy and convulsions, encephalopathy and delirium, and parkinsonism. A study was conducted on 328 children with Japanese encephalitis to determine the risk factors of postencephalitic epilepsy and construct a risk prediction model. In the study, 65.9% of the children had seizures in the acute phase, and 14.7% developed postencephalitic epilepsy. Seizure number greater than 5, status epilepticus, and coma were identified as independent risk factors for postencephalitic epilepsy (16).
A retrospective, multicenter study of 1626 patients with Japanese encephalitis from the Chinese Center for Disease Control and Prevention (2016–2020) identified 230 (14%) with peripheral nerve injury (105). These patients with peripheral nerve injury, primarily in northwest China, exhibited acute flaccid paralysis and severe respiratory muscle paralysis necessitating longer hospital and intensive care unit stays. The mortality rate was higher in these patients (16%) compared to patients without peripheral nerve injury (5%).
Japanese encephalitis is a risk for travelers who visit Japanese encephalitis-endemic countries. Hills and colleagues reviewed all published Japanese encephalitis cases in travelers from nonendemic areas from 1973 through 2008 and assessed factors related to risk of infection. The authors collected 55 cases that occurred in citizens of 17 countries. Ten (18%) persons died, and 24 (44%) had mild to severe sequelae (38).
A 13-year-old Vietnamese boy presented with progressive weakness of the lower limbs following an initial febrile illness. During the first 2 days, he experienced fever, headache, vomiting, and neck stiffness. By day 3, he developed painful weakness in the left leg, which rapidly progressed to the right leg and lumbar spine. By day 5, he was unable to walk or sit without support. There was no significant past medical or family history, and he had received full immunization for diphtheria, tetanus, and polio. On examination, he was febrile (40.3°C), with tachycardia (130 bpm) and tachypnea (38 breaths/min). He was conscious with neck stiffness but no cranial nerve or upper limb involvement. The lower limbs showed marked flaccid paralysis (Medical Research Council grade 0/5 except 1/5 at ankles and great toes), absent deep tendon, abdominal, and cremasteric reflexes, with preserved sensation. Blood smear was negative for malaria; hematology showed leukocytosis (16,000/mm³, 69% neutrophils). Lumbar puncture revealed mildly elevated opening pressure, two white cells/mm³, 84 red cells/mm³, raised protein (74 mg/dL), and normal glucose levels. Gram stain, culture, chest x-ray, Mantoux test, and enterovirus stool culture were negative. Immunoassays detected Japanese encephalitis virus-specific IgM in both cerebrospinal fluid and serum (95), confirmed by ELISA with levels of 186 units (CSF) and 153 units (serum) (normal < 40). On day 7, he became confused but improved by day 10. At 2 months, he could walk with difficulty, and at 1-year follow-up, he had persistent gait abnormality and right leg muscle wasting (93).
Japanese encephalitis is caused by the Japanese encephalitis virus, a member of the flavivirus family. The virus has a small lipoprotein envelope measuring approximately 50 nanometers, which encloses a nucleocapsid made up of a core protein and a single-stranded RNA genome approximately 11 kilobases in length, with a molecular weight of 3800 kilodaltons. There are at least five genotypes of the virus circulating in Asia (19; 18; 101). Other neurotropic flaviviruses with similar epidemiological and clinical profiles exist worldwide, including Murray Valley encephalitis virus in Australia and St. Louis encephalitis virus in the United States (90; 88). West Nile virus, which was once limited to Africa, Europe, and the Middle East, entered North America in 1999 and has since caused widespread outbreaks (10; 77).
The genome of Japanese encephalitis virus includes untranslated regions at both ends and codes for seven nonstructural and three structural proteins. The envelope protein is a major structural component of the virus’s surface projections and plays a vital role in viral attachment, membrane fusion, and cell entry (74). Heavily sulphated heparan sulphate has been identified as a potential receptor for flavivirus entry (20). In animal models, the envelope protein appears to influence virulence, but it is uncertain whether this affects clinical manifestations in humans.
In endemic regions, nearly all children are infected with Japanese encephalitis virus, yet only 1 in 25 to 1 in 1000 develops clinical illness (102). The reasons behind this variation in disease manifestation remain unclear. Possible contributing factors include viral characteristics, such as virulence, route of entry, and viral load, as well as host-specific factors like age, immune status, genetic predisposition, and overall health. One study reported that polymorphisms in the TLR3 gene might influence genetic susceptibility to Japanese encephalitis among individuals in India (08).
Following a mosquito bite, the virus initially replicates locally and in regional lymph nodes, leading to a transient viremia. It eventually reaches the central nervous system through hematogenous spread. Once inside neurons, the virus replicates within the rough endoplasmic reticulum and Golgi apparatus, ultimately destroying these structures (100). Experimental studies indicate that viral infection disrupts the permeability of the blood-brain barrier, allowing virus particles or infected leukocytes to enter the brain, where infection typically begins (64).
Autopsy findings in patients who succumb to Japanese encephalitis reflect an extensive inflammatory response to neuronal infection. Brain regions most affected include the thalamus, basal ganglia, midbrain, cerebellum, and anterior horns of the spinal cord. These pathological findings correlate with clinical features, such as tremors, dystonia, and flaccid paralysis. Common histopathological findings include glial nodules, necrotic foci, and involvement of white matter (72; 48; 33). Perivascular cuffing, parenchymal infiltration by inflammatory cells, and phagocytosis of infected neurons are often observed. In cases in which death occurs rapidly, inflammation may not be evident histologically, though immunohistochemistry can detect viral antigen in apparently normal neurons (48). A study comparing human brain tissue and a mouse model identified edema, hemorrhage, perivascular inflammation, microglial nodules, and necrotic foci, with additional evidence of viral replication within vascular endothelium (29).
Both humoral and cellular immune responses play essential roles in the body’s defense against Japanese encephalitis. The humoral response, particularly during primary infection, is characterized by a strong IgM response in both serum and cerebrospinal fluid. By day 7 of illness, most patients demonstrate high IgM titers, and virus isolation is usually unsuccessful at this point (11). The absence of an IgM response correlates with positive virus isolation and fatal outcomes (58; 60). Although asymptomatic infection leads to elevated IgM in serum, cerebrospinal fluid IgM is not present (11). In individuals with secondary flavivirus infections—due to previous dengue infection or yellow fever vaccination—there is an anamnestic IgG response followed by a delayed IgM rise (11).
The cellular immune response is believed to restrict viral replication before central nervous system invasion, as demonstrated in animal models. In humans, specific T-cell responses have been identified in convalescent patients and vaccine recipients. These include CD8+ cytotoxic T-lymphocyte proliferation and CD4+ helper T-cell responses that recognize the envelope protein in a human leukocyte antigen-restricted manner (01). T-cells from individuals with prior subclinical infection show responses to multiple viral proteins, including PrM, E, NS1, NS3, and NS5 (52). Responses to the NS3 protein were particularly prominent, with both CD4+ and CD8+ T-cells producing interferon-gamma in a manner consistent with a T-helper 1 immune profile (51). Interestingly, patients evaluated 6 months after discharge showed similar proliferative responses to exposed healthy individuals, though they had reduced interferon-gamma production as measured by enzyme-linked immunosorbent assay (50).
Microglial activation appears to be a key contributor to disease pathogenesis. This activation may promote bystander neuronal injury following infection with Japanese encephalitis virus (30). Experimental models indicate that neuronal death through apoptosis, as well as the activation of microglia and astrocytes, are central to the disease process. Infected neurons release cytokines that activate surrounding glial cells, which, in turn, release inflammatory and pro-apoptotic mediators. This leads to further neuronal apoptosis, including uninfected bystander neurons. The activation of astrocytes, microglia, and endothelial cells facilitates recruitment of immune cells and contributes to blood-brain barrier breakdown (75). Elevated levels of proinflammatory cytokines and chemokines in the cerebrospinal fluid are associated with worse clinical outcomes in patients with Japanese encephalitis (106).
The Japanese encephalitis virus belongs to the same family as the viruses responsible for dengue, yellow fever, Zika, and West Nile fever. It is the leading cause of viral encephalitis in many parts of Asia, with an estimated 100,000 symptomatic cases and around 25,000 deaths reported annually. Most infections are mild or without symptoms, but about one in 250 individuals develops severe disease. The death rate among those with symptoms can be as high as 30%, and up to half of survivors may suffer permanent neurologic, cognitive, or behavioral problems. The infection is mainly seen in children under 15 years of age and spreads through the bite of infected mosquitoes, especially in rural or farming areas near pigs and water birds. Recent shifts in Japanese encephalitis epidemiology include a rising number of adult cases due to expanded transmission zones, vaccination in children reducing childhood infections, and increased travel to endemic regions. The clinical features are similar across age groups, although seizures are less common in adults. Strengthening childhood vaccination in endemic areas remains essential to lessen disease burden in both children and adults (39).
Japanese encephalitis exhibits distinct geographical and seasonal patterns across Asia. In northern regions, such as northern Vietnam, northern Thailand, Korea, Japan, Taiwan, China, Nepal, and northern India, large-scale epidemics occur during the summer months. In contrast, southern areas like southern Vietnam, southern Thailand, Indonesia, Malaysia, the Philippines, Sri Lanka, and southern India experience year-round endemic transmission with a peak following the onset of the rainy season (102). These regional differences are likely influenced by climatic conditions and mosquito abundance, although viral genotype and host susceptibility may also play roles.
Japan has monitored Japanese encephalitis since 1965 through the National Epidemiological Surveillance of Vaccine Preventable Diseases. In the late 1960s, over 1000 cases were reported annually, but the number drastically declined to fewer than 10 per year from 1992 to 2004. Between 1982 and 2004, 361 cases were reported, with prognosis data available for 320. Among them, 58 (18%) died, 160 (50%) had neuropsychiatric sequelae, and 102 (32%) recovered completely. Notably, 78% of these patients were aged 40 years or older, with the highest incidence in the 60 to 69 age group. The majority of cases occurred in unvaccinated individuals (03).
A report by the Centers for Disease Control and Prevention (2009) described three cases among travelers from the United States who had visited Asia. These individuals were Asian immigrants or relatives, all unvaccinated. Each presented with fever and altered mental status; however, only one was diagnosed with Japanese encephalitis early in the disease course. Although all patients recovered, two were left with neurologic deficits.
Serological testing of cerebrospinal fluid in an endemic region found that Japanese encephalitis accounted for 26.9%, 9.9%, and 14.8% of acute encephalitic syndrome cases in 2011, 2012, and 2013, respectively. Among confirmed cases, 30% were adults, and males were more frequently affected. The outbreak period coincided with the monsoon and post-monsoon seasons (46). In Uttar Pradesh, a northern Indian state, 47,509 cases of acute encephalitis syndrome were recorded between 2005 and 2018. Of these, approximately 10% had confirmed Japanese encephalitis (86).
The Japanese encephalitis virus is transmitted by Culex mosquitoes, particularly Culex tritaeniorhynchus, in an enzootic cycle involving pigs, chickens, and other birds. The mosquito typically breeds in stagnant water, such as rice fields (45). Humans, who acquire the virus incidentally through proximity to the enzootic cycle, are considered dead-end hosts due to brief, low-level viremia. In rural Asia, most people are infected in childhood or early adulthood, with about 10% of the susceptible population infected annually (40). Most human infections are asymptomatic or result in mild, flu-like illness.
Over the past 50 years, the geographical range of the virus has expanded. Epidemics have occurred in China since 1935, in South Korea from the late 1940s, and in Vietnam and Thailand since the mid-1960s (102). The disease was identified in southern India in 1955 and spread northward from the 1970s onward. Its movement into Nepal suggests further northwestern expansion (02). Outbreaks have extended across Southeast Asia and the Pacific Rim, reaching mainland Australia in 1998 (35). The factors driving this spread remain unclear but may include changes in agricultural practices like increased irrigation and animal farming, which favor mosquito breeding and vertebrate host availability. In developed countries, such as Japan, Taiwan, and South Korea, mass childhood vaccination and socioeconomic development have reduced case numbers (90). However, vaccination in children in South Korea has coincided with a relative increase in adult cases (102).
A notable outbreak occurred in Australia, with 40 confirmed or probable cases and five deaths. A serosurvey conducted during the 2022 outbreak in five towns in New South Wales revealed that 8.7% of 917 participants had antibodies to the Japanese encephalitis virus (05). Seropositivity was highest in those aged 65 and older (13.7%) and was more common in males (10.6%) than in females (7.5%). This outbreak, the first since 1998, involved genotype 4 strains related to those in Indonesia. Its emergence was likely facilitated by migratory birds, mosquitoes, or both, with pigs acting as amplifying hosts. Contributing factors included La Niña weather, intensive pig farming, and a large feral pig population. Culex annulirostris was identified as the primary mosquito vector (103; 111).
Control strategies for Japanese encephalitis target both disruption of the virus's enzootic cycle and prevention of human infection, including vaccination. Although efforts such as Culex mosquito control and pig vaccination have been implemented, these measures have largely been ineffective. Preventing mosquito bites remains critical, particularly for travelers to endemic areas. Recommendations include minimizing exposed skin, using insect repellents containing DEET (N,N-diethyl-3-methylbenzamide), and adopting personal protective strategies, such as wearing long-sleeved clothing and using coils and vaporizers (110). Although effective for short-term visitors, these approaches may not be practical for long-term residents in endemic regions.
Historically, the most widely used vaccine has been a mouse brain–derived inactivated vaccine based on the Nakayama or Beijing-1 strain, developed in Japan. Its efficacy was demonstrated in large-scale, double-blind, randomized controlled trials in Taiwan and Thailand involving over 300,000 children (40; 84). This vaccine has a moderate incidence of mild side effects. Serious neurologic adverse events, such as encephalitis, encephalopathy, seizures, or peripheral neuropathy, are extremely rare, occurring in approximately one per million recipients. Since 1989, allergic reactions—such as itching, urticaria, and occasionally angioedema—have been noted, especially in individuals with preexisting allergies; these may occasionally require hospitalization and corticosteroid therapy (78). Cell culture–derived vaccines have largely replaced the older mouse brain–derived versions (109).
Since the late 1980s, a live attenuated vaccine cultured in baby hamster kidney cells has been safely administered to over 100 million children in China. It showed 97.5% efficacy with two doses given a year apart (37), and a single dose demonstrated protective efficacy even when administered shortly before exposure (07). Although licensed in China in 1988 and used in several Asian countries, it remains unavailable in Western nations. A novel chimeric vaccine based on the yellow fever 17D platform has also shown promising safety and immunogenicity (04; 73). However, a Cochrane review concluded that because only one of three vaccines has been tested in randomized controlled trials, comparing their relative efficacy remains difficult (84).
|
|
Live attenuated vaccine (SA 14-14-2 strain): | |
|
• First dose is given subcutaneously at 8 months of age, followed by a booster dose at 2 years of age. | ||
|
|
Inactivated, Vero cell-derived, alum-adjuvanted vaccine (SA 14-14-2 strain): | |
|
• Primary immunization consists of two intramuscular doses, 4 weeks apart. A booster is recommended after year 1. | ||
|
|
Inactivated, Vero cell-derived vaccines (Beijing-1 strain): | |
|
• Primary immunization consists of three doses at days 0, 7, and 28, or two doses given preferably 4 weeks apart (0.25 ml for children < 3 years, 0.5 ml for all other ages). One booster is recommended 12 to 14 months after completion of the primary immunization, and thereafter every 3 years. | ||
|
|
Live chimeric vaccine (with yellow fever 17D as backbone): | |
|
• A single dose is recommended. | ||
|
| ||
In a multicenter randomized controlled trial, Li and colleagues demonstrated that co-administration of the live-attenuated Japanese encephalitis vaccine with the measles and rubella vaccine does not compromise the seropositivity rates of measles and rubella, supporting the simultaneous use of both vaccines in Japanese encephalitis–endemic countries (59). Japanese encephalitis vaccination is recommended for international travelers planning to stay for 1 month or more in endemic areas during the transmission season, as well as for laboratory personnel at risk of exposure to infectious Japanese encephalitis virus (27). Due to the unpredictable nature of Japanese encephalitis outbreaks, their variability from year to year, and limitations in epidemiological data, identifying high-risk regions is difficult, prompting calls for broader vaccine implementation (45; 13).
A newer purified, inactivated Japanese encephalitis vaccine (IC51), produced using Vero cell cultures, has been shown to be both safe and immunogenic and is currently recommended for international travelers (47). Surveillance data from the U.S. Vaccine Adverse Event Reporting System covering 1999 to 2009 revealed 300 adverse event reports following the use of inactivated mouse brain–derived Japanese encephalitis vaccine (24 per 100,000 doses), including 106 (35%) hypersensitivity reactions and four (1%) neurologic events (62). Serious adverse events were reported in only 23 cases (1.8 per 100,000 doses).
A Chinese study suggested a potential association between the universal use of live-attenuated Japanese encephalitis vaccine and increased childhood-onset myasthenia gravis, possibly due to autoimmune responses triggered by molecular mimicry involving acetylcholine receptors (36). A One Health strategy—incorporating human, animal, vector, and environmental surveillance—is essential for early detection and response to Japanese encephalitis virus activity. This integrated approach is vital for guiding public health action, especially as the virus’s expanding presence, including in Australia, raises concerns for its spread to new regions (67).
Infection with Japanese encephalitis virus may be asymptomatic or may cause febrile illness, meningitis, myelitis, or encephalitis. Encephalitis is the most commonly recognized presentation and is clinically indistinguishable from other causes of an acute encephalitis syndrome. Clinically, a case of acute encephalitis syndrome is defined as a person of any age, at any time of year, with the acute onset of fever and a change in mental status (including symptoms such as confusion, disorientation, coma, or inability to talk) and new onset of seizures (excluding simple febrile seizures). Other early clinical findings may include an increase in irritability, somnolence, or abnormal behavior greater than that seen with usual febrile illness (108). Lymphocytic CSF with an elevated protein and a normal glucose ratio supports the diagnosis, but other causes must be excluded, and virological confirmation is needed. The differential diagnosis is broad, particularly in adults, and can be divided into viral encephalitides, nonviral CNS infections, infectious diseases with CNS manifestations, and noninfectious diseases:
|
Viral encephalitides | |
|
• Arboviruses | |
|
Nonviral CNS infections | |
|
• Acute bacterial meningitis | |
|
Infectious diseases with CNS manifestations | |
|
• Febrile convulsions | |
|
Noninfectious diseases | |
|
• Reye syndrome | |
Japanese encephalitis can be mistaken for other arboviral encephalitides, especially in regions where multiple neurotropic viruses are endemic. Illnesses caused by other flaviviruses, such as Murray Valley encephalitis, West Nile virus, and even dengue virus, may present with similar clinical features (44; 92). Additionally, outbreaks of Nipah virus encephalitis (22), enterovirus 71 (107), and Chandipura virus (80) have highlighted the importance of not hastily attributing all encephalitis cases to Japanese encephalitis virus. Differentiating Japanese encephalitis from conditions such as partially treated bacterial meningitis or cerebral malaria is often challenging, particularly in tropical regions where antibiotics and antimalarials are easily accessible without prescription. Based on their findings in an endemic area, Dubot-Pérès and colleagues cautioned that detection of anti-Japanese encephalitis virus IgM in cerebrospinal fluid may be misleading and stressed the importance of excluding other treatable infections, especially bacterial, before confirming the diagnosis of Japanese encephalitis. In their study, 6% of Japanese encephalitis IgM-positive CSF samples were found to be associated with other pathogens (26).
Garg and colleagues reported a unique case of acute encephalitis syndrome in which IgM antibodies against both dengue virus and Japanese encephalitis virus were detected in serum and CSF. In regions endemic for both viruses, such co-positivity may result from either true co-infection or from cross-reactivity of serological tests, a known phenomenon between flaviviruses (28). Another report described a similar case with MRI evidence of bilateral thalamic and brainstem lesions and dual seropositivity (87).
Acute flaccid paralysis caused by Japanese encephalitis virus must be distinguished from other anterior horn cell infections, including poliovirus, non-polio enteroviruses, and West Nile virus, as well as from post-infectious conditions like Guillain-Barré syndrome. Although Japanese encephalitis–associated myelitis may resemble poliomyelitis, concurrent bladder involvement favors Japanese encephalitis (94). Enterovirus 71 is a notable cause of flaccid paralysis, particularly during hand-foot-and-mouth disease outbreaks (69; 42). Differentiating Japanese encephalitis from the acute inflammatory demyelinating polyneuropathy form of Guillain-Barré syndrome is generally straightforward but more difficult with the acute motor axonal neuropathy variant, which also presents with lower motor neuron involvement and has been reported in large Asian epidemics (68). However, Guillain-Barré syndrome is usually symmetrical and post-infectious, unlike Japanese encephalitis, which occurs during febrile illness.
Japanese encephalitis virus may also trigger autoimmune responses. Like herpes simplex virus, it can induce anti-N-methyl-D-aspartate receptor (NMDAR) antibody production, resulting in autoimmune encephalitis (21; 99). In a study by Ma and colleagues, five of 63 patients with Japanese encephalitis relapsed during recovery; three were diagnosed with anti-NMDAR encephalitis, presenting with choreoathetosis or psychiatric symptoms (65). Liu and colleagues studied 31 children with Japanese encephalitis and found that five relapsed: two developed anti-NMDAR antibodies, one developed anti-GABA-B receptor antibodies, and two produced antibodies against unknown neuronal membrane antigens (63). Although the precise mechanism remains unclear, synaptic autoimmunity may arise from virus-induced neuronal antigen exposure.
Coinfection of neurocysticercosis with Japanese encephalitis is also commonly seen in endemic regions and has been associated with poorer outcomes (34; 98).
In most patients with Japanese encephalitis, there is a peripheral neutrophil leukocytosis. Typically, there is a moderate CSF pleocytosis of 10 to 100 cells per mm3, with predominant lymphocytes, mildly elevated protein (50 to 200 mg%) and a normal glucose ratio. However, polymorphonuclear cells may predominate early in the disease, or there may be no CSF pleocytosis.
Computed tomography may show bilateral nonenhancing low-density areas in the thalamus, basal ganglia, midbrain, pons, and medulla (71). The most consistent MRI findings in Japanese encephalitis are bilateral thalamic lesions with or without hemorrhagic changes. Lesions are also noted in the substantia nigra, brainstem, cerebellum, cerebral cortex, and white matter. Lesions in the substantia nigra may be characteristic of patients with parkinsonian features (79). On MRI, there may be more extensive changes and characteristic thalamic lesions of mixed intensity on T1 and T2; weighted scans are suggestive of hemorrhage (57). These changes may be useful in distinguishing Japanese encephalitis from Herpes simplex encephalitis, where the changes are characteristically frontotemporal. Medial temporal MR lesions have been reported in several other diseases of central nervous system-like paraneoplastic limbic encephalitis and neurosyphilis.
In Japanese encephalitis, medial temporal lesions are often associated with bilateral thalamic or midbrain involvement. Temporal lobe involvement in Japanese encephalitis dominantly affects the hippocampus (33; 49).
A variety of electroencephalographic abnormalities have been reported in Japanese encephalitis including theta and delta coma, burst suppression, epileptiform activity, and occasionally alpha coma, but none seem to be diagnostic.
Specific tests for diagnosing Japanese encephalitis demonstrate either the presence of the virus or the antibody to the virus. Attempts to isolate Japanese encephalitis virus from clinical specimens are usually unsuccessful because of low titers and the rapid production of neutralizing antibody. Isolates are sometimes obtained from CSF (in which case it is associated with a failure of antibody production and a high mortality rate) (58) or from brain tissue (either at autopsy or post-mortem needle biopsy). Japanese encephalitis virus RNA has been detected in human CSF samples using reverse transcriptase polymerase chain reaction (44). However, its reliability as a routine diagnostic test has yet to be shown. Of the various serological methods available, the IgM capture ELISA has proved the most useful. The presence of anti-Japanese encephalitis virus IgM in the CSF has a sensitivity and specificity of less than 95% for CNS infection with Japanese encephalitis virus (11). The ELISA was modified to a simple “dot-blot” nitrocellulose membrane-based format for diagnosis of Japanese encephalitis for use in the rural settings where the disease often occurs (95).
World Health Organization recommended laboratory criteria for confirmation (108):
|
(1) Presence of Japanese encephalitis virus-specific IgM antibody in a single sample of cerebrospinal fluid or serum, an IgM-capture ELISA specifically for Japanese encephalitis virus. | |
|
(2) Detection of Japanese encephalitis virus antigens in tissue by immunohistochemistry. | |
|
(3) Detection of Japanese encephalitis virus genome in serum, plasma, blood, cerebrospinal fluid, or tissue by reverse transcriptase polymerase chain reaction or an equally sensitive and specific nucleic acid amplification test. | |
|
(4) Isolation of Japanese encephalitis virus in serum, plasma, blood, cerebrospinal fluid, or tissue. | |
|
(5) Detection of a 4-fold or greater rise in Japanese encephalitis virus-specific antibody as measured by hemagglutination inhibition or plaque reduction neutralization assay in serum collected during the acute and convalescent phase of illness. |
Currently, there is no specific antiviral therapy available for Japanese encephalitis. Although several compounds, such as nitric oxide, ribavirin, and interferon alpha, have demonstrated efficacy in vitro or in animal studies, clinical results have been largely disappointing (83; 96). A randomized controlled trial evaluating oral ribavirin in patients with Japanese encephalitis found it to be ineffective (56). Similarly, interferon alpha-2a was tested in a double-blind, placebo-controlled trial involving children but yielded negative outcomes (83; 96; 31).
Minocycline showed promise in experimental models, providing complete protection in mice. Treatment led to reduced neuronal apoptosis, microglial activation, caspase activity, inflammatory mediators, and viral titers (70). However, a randomized trial involving pediatric patients with acute encephalitis syndrome, including those with Japanese encephalitis, did not show significant clinical benefit (53).
Intravenous immunoglobulin (IVIG) has also been explored. In a trial conducted in Nepal, IVIG was found to enhance neutralizing antibody production, suggesting its therapeutic potential, though further studies are needed (81). A study in China involving 124 children with severe Japanese encephalitis demonstrated that IVIG use was associated with shorter durations of unconsciousness, seizures, and neurologic signs, such as light reflex dullness and pyramidal and meningeal signs (15). In Korea, a small series of eight patients treated with IVIG combined with ribavirin and interferon alpha 2b showed partial or complete recovery in five patients, compared to poorer outcomes in patients who received only supportive care (97).
Supportive treatment remains the cornerstone of care. Management includes controlling seizures and raised intracranial pressure. Despite earlier use, corticosteroids like dexamethasone were found to be ineffective in a randomized trial (41). Preventive care with physiotherapy and nursing is essential to avoid complications, such as aspiration pneumonia, bedsores, malnutrition, and contractures.
Japanese encephalitis occurred in five pregnant women during an epidemic in India, two of whom aborted and one of who had an apparently normal child. The virus was isolated from one of the aborted fetuses (14).
All contributors' financial relationships have been reviewed and mitigated to ensure that this and every other article is free from commercial bias.

Ravindra Kumar Garg DM
Dr. Garg of King George's Medical University in Lucknow, India, has no relevant financial relationships to disclose.
See Profile
John E Greenlee MD
Dr. Greenlee of the University of Utah School of Medicine has no relevant financial relationships to disclose.
See ProfileNearly 3,000 illustrations, including video clips of neurologic disorders.
Every article is reviewed by our esteemed Editorial Board for accuracy and currency.
Full spectrum of neurology in 1,200 comprehensive articles.
Listen to MedLink on the go with Audio versions of each article.
MedLink, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Infectious Disorders
May. 15, 2026
General Neurology
May. 13, 2026
Infectious Disorders
May. 12, 2026
Infectious Disorders
May. 12, 2026
Infectious Disorders
May. 05, 2026
Infectious Disorders
May. 01, 2026
Infectious Disorders
Apr. 30, 2026
Headache & Pain
Apr. 10, 2026