




























































































































Developmental Malformations
Vein of Galen malformations
Sep. 22, 2024
MedLink®, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Worddefinition
At vero eos et accusamus et iusto odio dignissimos ducimus qui blanditiis praesentium voluptatum deleniti atque corrupti quos dolores et quas.
Anencephaly is a severe and uniformly lethal malformation that results from incomplete closure of the anterior neural tube, whereby fetuses or infants present with absent calvarial and cerebral structures. In this article, the author discusses the etiology, pathogenesis, genetic contribution, and epidemiology of anencephaly as well as approaches toward the diagnosis and prevention of this and other neural tube defects.
|
• Anencephaly represents one of the major forms of neural tube defect, along with encephalocele and spina bifida. | |
|
• The prenatal administration of folic acid is essential in preventing neural tube defects; other vitamins and dietary substances (eg, inositol) may exert a preventative effect as well. | |
|
• Some cases of anencephaly occur from mechanisms that do not involve folic acid (so-called "folate-resistant" neural tube defects, although a more accurate term would be “folate nonresponsive”). |
Anencephaly was not described in recognizable form until the 16th century (16; 189). In the mid-17th century, various pamphlets were published concerning monstrous births, and the descriptions and images suggest anencephaly (11). Some of the earliest clinical photographs of anencephaly were taken in the late 1860s and published in the Revue photographique des hôpitaux de Paris.
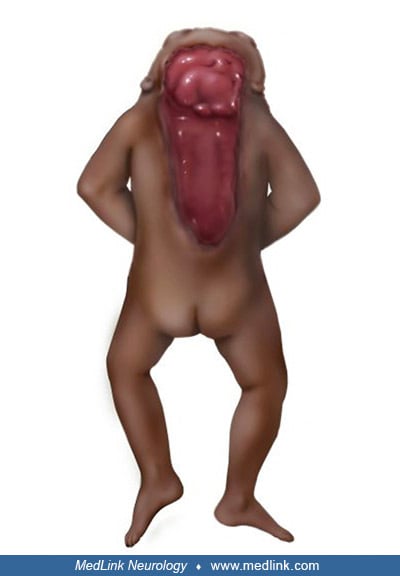
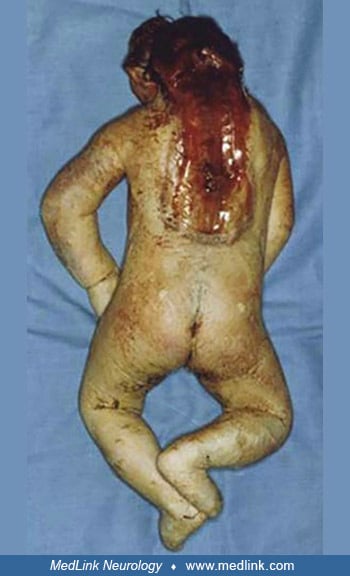
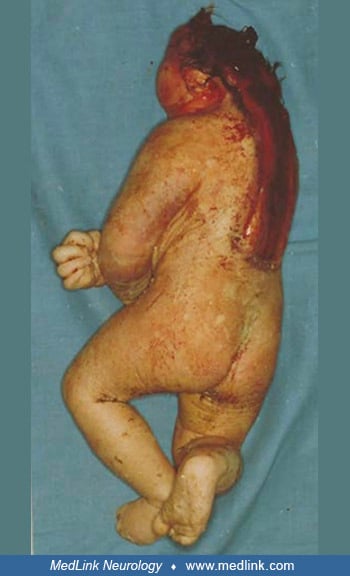
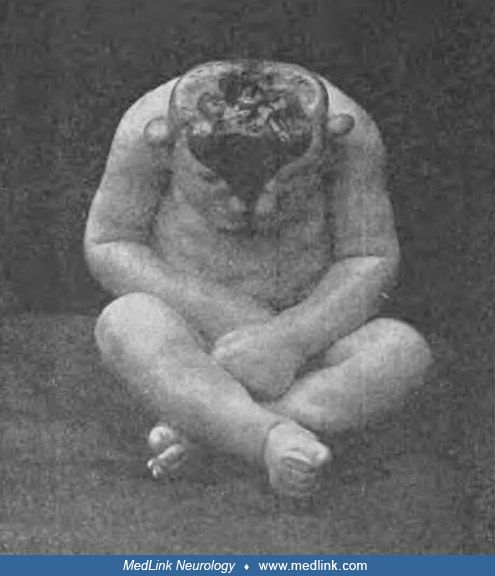
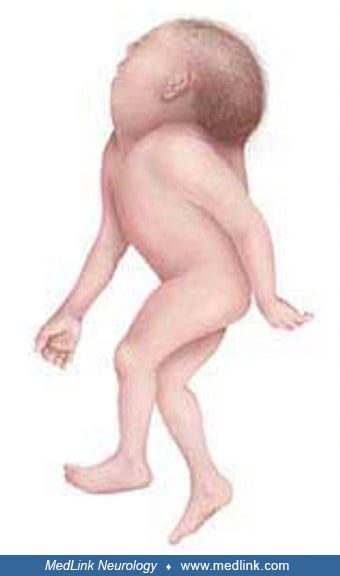
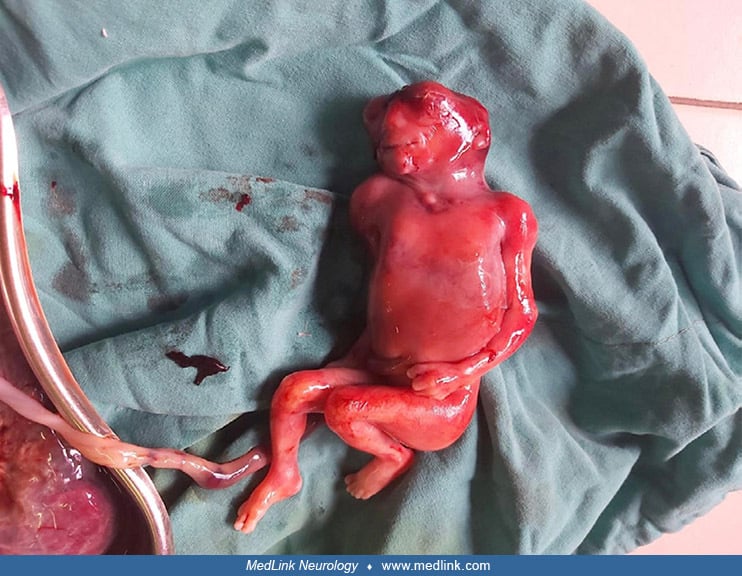
Anencephaly belongs to a group of congenital malformations known collectively as "neural tube defects" (which also include encephalocele and myelomeningocele). The common neuroanatomical feature in anencephaly is an open defect in the calvaria and skin. The defect varies in size but is often so large that the predominant feature is a degenerated and hemorrhagic mass of tissue lying on an essentially exposed basicranium. Numerous classifications and synonyms have led to confusion about the spectrum of anencephaly.
Terminology of anencephaly and similar or related disorders. Acephalus, acrania, anencephalus, cranioschisis, and cranial or cerebral dysraphia are commonly used but not necessarily correctly.
Acephaly is the congenital absence of a head.
The term "acrania," literally meaning the absence of the cranium, is used to refer to a rare congenital anomaly in which the flat bones of the cranial vault are partially or completely absent, although the bones at the base of the skull are normally present, often with abnormal development of the cerebral hemispheres. The condition is frequently, though not always, associated with anencephaly.
The term "acalvaria" is often taken as a synonym for acrania. According to the Orphanet definition, primary acalvaria is defined as follows (ORPHA:945):
|
A rare malformation characterized by missing scalp and flat bones over an area of the cranial vault. The size of the affected area is variable. In rare cases, acalvaria involves the whole of the dome-like superior portion of the cranium comprising the frontal, parietal, and occipital bones. Dura mater and associated muscles are absent in the affected area but the central nervous system is usually unaffected, although some neuropathological abnormality is often present (eg, holoprosencephaly or gyration anomalies). Skull base and facial bones are normal. |
Neural tube defects are malformations of the brain and spinal cord that occur when the neural tube does not close properly. These types of birth defects develop very early during pregnancy, often before a woman knows she is pregnant. Neural tube defects include forms of anencephaly (holoanencephaly, meroanencephaly, and craniorachischisis), iniencephaly, encephalocele, and forms of spina bifida (myelomeningocele, meningocele). The most common neural tube defects are anencephaly and spina bifida.
Anencephaly, or anencephalus, is a severe form of anterior neural tube defect in which the brain is absent.
Isolated anencephaly/exencephaly is defined as follows (ORPHA:1048):
|
This malformation is characterized by the total or partial absence of the cranial vault and the covering skin, the brain being missing or reduced to a small mass. Most cases are stillborn, although some infants have been reported to survive for a few hours or even a few days. ... Anencephaly is likely to be multifactorial, the result of gene-environment interactions. ... Familial cases with a seemingly autosomal recessive mode of inheritance have been described but most cases are sporadic. |
Cranioschisis is a congenital abnormality characterized by the failure of the bones of the skull to close, whereas rachischisis is a neural tube defect in which an extended portion of the spinal cord remains open. These often occur together as craniorachischisis, a neural tube defect in which both the brain and spinal cord remain open to varying degrees.
One classification that provides a framework for all types of anencephaly differentiates craniorachischisis (head and spine are open) from holoacrania (cranial defect extends through the foramen magnum) and meroacrania (partial cranial defect not extending to foramen magnum). However, the latter two disorders are most often referred to in terms of the brain structures involved (ie, holoanencephaly and meroanencephaly, respectively) rather than solely in terms of the extent of bony abnormality.
Cranial dysraphism collectively refers to anterior neural tube defects, anencephaly, and encephaloceles that result from failure of closure of the anterior end of the neural tube. In contrast, spinal dysraphism, or spina bifida, refers to posterior neural tube defects that result from failure of closure of the caudal end of the neural tube.
Exencephaly is a malformation of the neural tube with a large amount of protruding brain tissue and absence of calvarium. Animal studies generally use the term "exencephaly" to refer to a disorder equivalent to anencephaly, and in the Orpha.net terminology database, they are synonyms.
Encephaloceles. According to the Centers for Disease Control and Prevention definition, an encephalocele is a neural tube defect characterized by a “pedunculated or sessile cystic, skin-covered lesion protruding through a defect in the cranium.” Encephaloceles can contain herniated meninges and brain tissue (encephalocele or meningoencephalocele) or only meninges (cranial meningocele). If a portion of the lateral ventricle extends extracranially, the defect is called a hydrencephalocele (hydrocephalocele, hydroencephalocele). Encephaloceles can vary in size and location. The most common type is occipital (74%), followed by parietal (13%), but less common types include frontal, nasofrontal, orbital, nasal, and extensive forms defying precise localization. Encephalocele is more likely to occur with other unrelated structural anomalies or syndromes than either anencephaly or spina bifida.
Spina bifida. Spina bifida, the most common type of neural tube defect, is a general term used to describe a defect in which part of the meninges, spinal cord, or both protrudes through an opening in the vertebral column. Hydrocephalus is a common complication, especially among children with open meningomyeloceles.
Specific types of spina bifida include:
|
Meningocele. Herniation of the meninges through a spinal defect, forming a cyst filled with cerebrospinal fluid. | |
|
Meningomyelocele (myelomeningocele). Protrusion of the meninges and spinal cord through an opening in the vertebral column. | |
|
Myelocele (myeloschisis). A splayed vertebral column and plaque-like spinal cord without membrane or skin covering. |
Spina bifida may occur at any level of the spinal cord: cervical, thoracic, lumbar, or sacral.
|
• Liveborn infants with anencephaly generally die quickly. | |
|
• Affected infants can have seemingly purposeful movements, startle myoclonus, increased tone and deep tendon reflexes, normal cardiovascular status, and regular respiratory patterns. | |
|
• Because of the severity of impairment and frequency of early death, many workers discuss the option of termination of pregnancy with families. |
There are three forms of anencephaly: holoanencephaly (total), meroanencephaly (partial), and craniorachischisis.
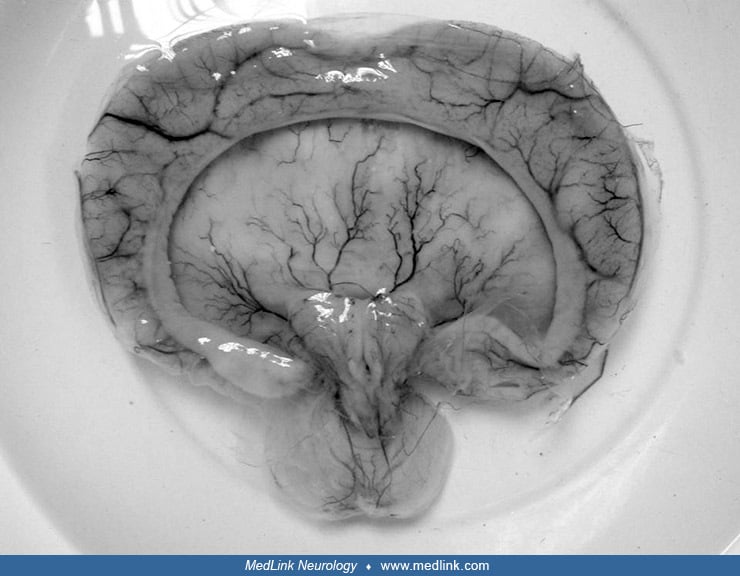
Wet preparation in the Anatomisches Museum der Universität, Basel, Switzerland. (Photograph by Mattes on October 5, 2012. Creative Commons Attribution 2.0 Germany License, https://creativecommons.org/licenses/by/2.0/de/deed.en...
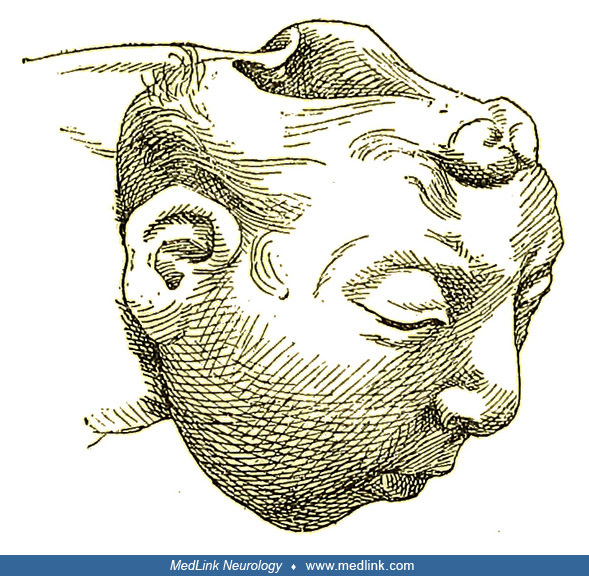
Although anencephaly is often an isolated, nonsyndromic anomaly, it not uncommonly manifests with other major anomalies (eg, cleft face, cleft palate, congenital heart defects). A retrospective analysis of 139 fetuses with acrania/exencephaly/anencephaly sequence diagnosed between 2006 and 2020 in a single tertiary referral ultrasound department found that (1) about a quarter of such fetuses have additional structural non-neural tube defect anomalies, which are mostly consistent with limb body wall complex; and (2) genetic abnormalities are diagnosed in almost 10% of affected fetuses, with trisomy 18 being the most common aberration (22). Factors that significantly increased the odds of genetic anomalies in fetuses with acrania/exencephaly/anencephaly sequence included male sex, limb anomalies, and ventral body wall defects. The eyes of anencephalics are normally formed; the apparent bulging of the eyes results from the absence of the frontal portion of the cranial vault. The cerebellum, brainstem, and spinal cord are generally intact.
Anencephaly most commonly involves singleton births, but it can occur with multiple births (eg, involving one or both of a set of twins) or, rarely, may involve one or both of a set of conjoined twins.
Lateral overview showing oblique positioning of the heads. The gluteal cleft of the left twin (black arrowhead) and the three lower extremities (black arrow) are distinguishable. Parapagus twins are conjoined twins that lie sid...
Detailed right-sided view of the genitalia showing two penises (black arrow), of which the lower-positioned one is hypoplastic. The right perineal raphe (black arrowhead) can be followed toward the anal opening (white arrowhead...
Three-dimensional computed tomographic reconstruction of the skeleton. Overview showing normally developed right twin’s cranium and anencephaly in the left twin. Two separate vertebral columns (purple arrow) and three upper (gr...
The "area cerebrovasculosa" of anencephalic fetuses is an irregular vascular proliferation resembling an angioma, within which are scattered primitive neuroglial tissue elements and disorganized neuroepithelial tissue. It typically appears as a bilaterally symmetrical cystic mass covering the anterior and middle cranial fossae, which is walled with glial tissue, partially lined with ependyma, and filled with abnormally vascular choroid plexus (96; 18; 157). The highly vascular covering of the cyst and the overlying squamous epithelium is in continuity with hairy skin at the margins of the lesion. The posterior cranial fossa contains a variable quantity of medulla oblongata that has no connection with the cystic mass. The degree of hindbrain development is associated with heterotopic glial tissue in the spinal subarachnoid space of some anencephalic fetuses (17; 18).
Liveborn infants with anencephaly generally die quickly, but some may live for days, weeks, or rarely, months. In an exceptional situation in 1995, an anencephalic infant, "Baby K," was kept alive on assisted ventilation by order of the Fourth Circuit Court of Appeals, a ruling that caused considerable professional duress (94; 286). To date, the longest survival of an unsupported infant has been 28 months (93). The prenatal ultrasound image published in this case suggested that more brain tissue was present than usual. Surprisingly, affected infants can have seemingly purposeful movements, startle myoclonus, increased tone and deep tendon reflexes, normal cardiovascular status, and regular respiratory patterns. They may have difficulty maintaining a normal body temperature and become hypothermic. Some infants with anencephaly make crying sounds and can swallow. Such lifelike activities create anxiety for caregivers. For these reasons, practitioners are cautioned in the use of careful and precise language as they counsel families, focusing on the specifics of prognosis rather than abstract generalities (371).
Because of the severity of impairment and frequency of early death, many workers discuss the option of termination of pregnancy with families (143). Some have suggested that organs of anencephalic infants might be used for transplantation; however, a significant proportion of these organs are often hypoplastic or malformed (189; 294; 221; 191). A medical task force on anencephaly reviewed the medical, social, legal, and ethical issues regarding anencephaly and recognized various complicating factors, including the difficulty in documenting brain death in affected infants (219). In 1995, the ethics council of the American Medical Association announced its support for procuring organs for transplantation from anencephalic infants; subsequent controversy caused the council to withdraw its opinion (367). This issue remains unresolved (261).
Approximately 65% of fetuses with anencephaly die in utero (219). Most liveborn infants with anencephaly die within minutes to hours of delivery, and nearly all die in the first week, though a few long-term survivors have been reported. A review of 26 patients showed that survival time ranged from 10 minutes to 8 days (249). Cases carried to term may be complicated by stillbirth, polyhydramnios (sometimes severe), cesarean delivery, shoulder dystocia, and possible obstetrical hemorrhage (98). In twin pregnancies discordant for anencephaly, the risk for premature delivery and low birth weight of the nonaffected twin is elevated, requiring increased monitoring and possible intervention (187; 198). In one small series, an increased risk for neural tube defect was observed among siblings of patients with lipomyelomeningocele (300).
A female infant was born at 39 weeks to a 22-year-old gravida 1 mother who suffered from cervical carcinoma. The prenatal course was unremarkable, and the mother gave no history of drug or alcohol use or smoking. The infant was delivered by cesarean section because of a breech presentation and given Apgar scores of 6 at one minute and 9 at five minutes. Moro-like reflexes and grasping could be elicited, but the infant manifested occasional seizure activity and choking. With increasing periodic apnea and bradycardia, she expired at 23 hours.
|
• Anencephaly is etiologically heterogeneous but is thought to involve an interaction between genetic and environmental factors. | |
|
• Known risk factors account for less than one half of cases. | |
|
• In mouse models of neural tube defects involving different mutations, some respond to folate (ie, with a reduced frequency of exencephaly or spina bifida aperta), whereas folate appears to have no preventative effect in others. | |
|
• In some folate-nonresponsive mutant mouse models of neural tube defects, other vitamins or dietary substances produce a preventative effect (eg, methionine, inositol, retinoic acid). | |
|
• Mouse mutants in folate pathway genes rarely result in neural tube defects. | |
|
• Folr1-null mouse embryos die by the tenth day of gestation with no neural tube closure. However, if they are treated with folinic acid, most homozygotes live to the 18th day of gestation, and 30% have exencephaly (among other defects of face, body wall, jaw, eyes, and limbs). Folr1 codes for folate receptor alpha, which suggests that a deficiency of folate entering the cells contributes to the failure of cranial neural tube closure. |
Anencephaly is etiologically heterogeneous (147; 188) but is thought to involve an interaction of genetic and environmental factors.
Micronutrient deficiencies of folate and vitamin B12 have been linked to an increased risk of neural tube defects, particularly inadequate material folate intake. Folate and vitamin B12 are metabolically linked through the folate and methionine cycles. Maternal folate intake is also potentially correctable.
At present, known risk factors account for less than one half of cases (02). Multiple etiologic factors have been implicated, but most have not been confirmed or are found only in isolated cases. Implicated factors include maternal exposure to certain drugs (eg, aminopterin, salicylates, clomiphene, and gonadotropins), infections (ie, influenza), nitrite-cured meats, blighted potatoes, and hard water. Difficulties in maternal glucose control (ie, diabetes), hyperinsulinemia, obesity, and intake of simple sugars have been associated with increased risk of neural tube defect and may act by affecting gene expression in the developing embryo (313; 290; 271; 353). Periconceptional maternal cannabis use is associated with the development of anencephaly (354; 278). Maternal hyperthermia, treatment of seizures in pregnant mothers with valproic acid, and the fungal product fumonisin are recognized causes of neural tube defects. One study identified an association of acute conditions (eg, maternal fever) and more chronic ones (eg, as migraine headaches) with neural tube defects (220). In another study, the greatest attributable risk for anencephaly was Hispanic ethnicity (02).
Overview potential underlying (patho)physiological pathways of folate deficiency and NTDs in obese women. Abbreviation: NTD = neural tube defect. (Source: van der Windt M, Schoenmakers S, van Rijn B, Galjaard S, Steegers-Theuni...
Genetic factors. A genetic basis for some cases of anencephaly seems likely, but it has been difficult to ascertain because the condition appears to be multigenic with complex gene-environment interactions (128). A genetic contribution has been suggested by the association with other conditions having a recognized genetic basis, such as anal stenosis, anterior sacral meningocele, and Meckel syndrome (92). A genetic basis is also suggested by the identification of numerous mutations whose function causes neural tube defects in mice (66; 128) and the occurrence of familial cases with a 10-fold increased risk among siblings (36). Familial cases can be striking, with recurrence of three to six affected infants per mother. Demenais and colleagues suggested that familial cases may involve monogenic inheritance with environmental influences (89), whereas Kalter and Warkany suggested that a genetic model would most likely involve multigenic inheritance (168; 169). Current thinking supports a multifactorial etiology (66; 128).
Analysis of sex ratios shows a predominance of female anencephalics, and many twins (usually discordant) have been reported. The increased incidence of anencephaly among twins does not appear to be related to the mode of conception (ie, natural vs. artificial) (20). Interestingly, this female predominance is more pronounced in those exhibiting localized cranial lesions than those with schisis involving the upper vertebral column (82).
A small percentage of individuals with anencephaly have an abnormal karyotype. In one study, 2% of affected fetuses had karyotype abnormalities, most often trisomy 18 or trisomy 13 (303). Two cases of anencephaly associated with partial duplication of 2p have been reported, suggesting that genes on the short arm of chromosome 2 could be important to CNS development (338).
A homozygous mutation in FOXN1, a member of the forkhead (or Fox) gene family, has been discovered in a human fetus with absent thymus, abnormal skin, anencephaly, and spina bifida (08). This finding suggests that the gene may be involved in neurulation in humans.
The presence of genetic factors is also supported by the observation that some 30% of defects are not prevented by folic acid supplements alone (25). It is possible that obesity (as quantified by basal metabolic index) influences the body distribution of folate (343; 353). The relationship of red blood cell folate levels and obesity appears to vary by basal metabolic index but requires further study (201).
Retinoic acid receptor signaling malfunctions have been observed in human neural tube defects, and the level of retinoic acid receptor gamma mRNA was significantly upregulated in the brain tissues of human fetuses with anencephaly (377).
Insufficiency of the ribosomal biogenesis gene treacle ribosome biogenesis factor 1 results in cranial neural tube defects and craniofacial malformations in mice (368). A p.(A491G) variant in TCOF1 is linked to anencephaly and single-nostril abnormality in the Han Chinese population. The pathogenic gene attenuates the production of total proteins, promotes nucleolar disruption, and stabilizes p53 protein, suggesting a loss-of-function mutation in ribosomal biogenesis and highlighting an unbalancing effect on cell apoptosis.
Mouse models. Mutant mouse models of neural tube defects provide insight into the mechanisms that result in neural tube defects (136; 138; 137; 139; 135; 355). Most of the (more than 50) single-gene mutations that cause neural tube defects in mice also cause lethal embryonic syndromes in which exencephaly is a nonspecific feature. Multigenic causes, such as those present in the curly-tail mutant mouse or the SELH/Bc mouse strain, produce nonsyndromic neural tube defects; 15% to 20% of the offspring of curly-tail mutant mice have spina bifida, whereas a similar percentage of the offspring of SELH/Bc mice have exencephaly (138; 355). "Spina bifida occulta" (ie, a dorsal gap in the vertebral arches overlying an intact neural tube) is genetically and developmentally unrelated to exencephaly or "spina bifida" (aperta). Almost all genetic forms of exencephaly or spina bifida aperta are caused by a failure of neural fold elevation (138). In addition, mutations causing neural tube defects affect distinct rostrocaudal zones along the neural folds through heterogeneous mechanisms; depending on which zone is affected, failure of neural fold elevation may variously produce "split face," exencephaly, rachischisis, or spina bifida (138; 256).
At the tissue level, failure of elevation may be caused by various processes (138):
|
• Slow growth of adjacent tethered tissue (eg, curly-tail mice) | |
|
• Defective forebrain mesenchyme | |
|
• A defective basal lamina in surface ectoderm | |
|
• Excessive breadth of the floorplate and notochord | |
|
• Abnormal neuroepithelium (eg, Sp or splotch mice) | |
|
• Morphological deformation of neural folds | |
|
• Abnormal neuroepithelial and neural crest cell gap-junction communication | |
|
• Incomplete compensation for a defective step in the elevation sequence (eg, the SELH/Bc mouse strain) |
At the biochemical and cellular levels, different mutants act through or affect different cellular processes (138):
|
• Faulty regulation of apoptosis (256) | |
|
• Premature differentiation | |
|
• Disruption of actin function | |
|
• An abnormal telomerase complex | |
|
• Faulty pyrimidine synthesis |
Normal (left) and exencephalic (right) fetal head in mouse model of congenital cranioschisis. Sagittal section of normal (left) and exencephalic (right) fetal head at embryonic day 17 (4x, hematoxylin and eosin). (Source: Oria ...
Cerebellum and brainstem in normal fetuses (left) and hypertrophic brainstem in exencephaly fetuses (right) (10x, hematoxylin and eosin). (Source: Oria M, Duru S, Figueira RL, et al. Cell necrosis, intrinsic apoptosis and senes...
Normal structured cerebral cortex (left) and brain cortex with no structures in exencephaly (right) (20x, hematoxylin and eosin). (Source: Oria M, Duru S, Figueira RL, et al. Cell necrosis, intrinsic apoptosis and senescence co...
Normal cortical layer organization in sham (top row) and loss of any cellular layer organization in exencephaly (bottom row) (left to right in each row: 10x, 20x, and 40x, hematoxylin and eosin). Abbreviations: MZ: marginal zon...
As the following examples illustrate, the neural tube defect preventative effect of maternal dietary supplementation is also heterogeneous, reflecting underlying differences in biochemical deficiencies and cellular processes.
|
• Maternal methionine supplementation, but not folinic acid or vitamin B12, prevents axial defects in Axd mouse neural tube mutants (105; 106; 138; 355). | |
|
• In curly-tail mice, inositol decreases neural tube defects by 55% to 85%, and retinoic acid decreases neural tube defects by 35% (134). Dietary methionine does not reduce penetrance of neural tube defects but causes a phenotype-specific decrease in embryonic growth (138; 355). | |
|
• Neural tube defects in the Splotch mutant can be "rescued" (ie, prevented) with either supplemental dietary folic acid or thymidine, suggesting that folic acid prevents neural tube defects by rescuing de novo thymidylate synthesis in this mouse model (375). |
Mouse mutants in folate pathway genes rarely result in neural tube defects. Folr1-null mouse embryos die by the tenth day of gestation with no neural tube closure, but if they are treated with folinic acid, most homozygotes live to the eighteenth day of gestation, and 30% have exencephaly (among other defects of face, body wall, jaw, eyes, and limbs). Folr1 codes for folate receptor alpha, which suggests that a deficiency of folate entering the cells contributes to failed cranial neural tube closure.
The curly-tail (ct) mutant mouse has been studied for more than 70 years and is now the best-understood mouse model of neural tube defect pathogenesis (301; 355). Expressions of ct gene mutations in curly-tail mice include prenatal death, spina bifida, and various degrees of kinky or curly tail. Failure of closure of the spinal neural tube in curly-tail mutant mice results in spina bifida, which is caused by a tissue-specific defect of cell proliferation in the tail bud of the E9.5 (ie, embryonic day 9.5) embryo. This cell proliferation defect results in a growth imbalance in the caudal region, leading to delayed neuropore closure and spina bifida, or tail defects. In addition to the principal ct gene on mouse chromosome 4, the curly-tail phenotype is influenced by several modifier genes and by environmental factors, illustrating a very clear, but complex, gene-environment interaction (355). Neural tube defects in curly-tail mice are "resistant" to correction by folic acid supplementation (as is the case in 30% of human neural tube defects), but they can be prevented by supplementation with myo-inositol (355). Curly-tail mice also exhibit enhanced susceptibility to the teratogenic effect of vitamin A given on day 8 of gestation, but if vitamin A is administered on day 9, the occurrence of neural tube defects is decreased rather than increased (302).
Similarly, human neural tube defects, including nonsyndromic anencephaly or spina bifida, may also reflect heterogeneous multigenic defects and complex gene-environment interactions that affect the developmental mechanisms responsible for elevation of the neural folds.
Embryology. The pathogenesis of anencephaly remains unclear. Two major schools exist: one holding that the primary mechanism is nonclosure of the rostral neural folds, and the other that cranial mesenchyme is primarily defective (204; 229). Most workers favor nonclosure of the neural tube as the primary mechanism, as evidenced by the widespread popularity of the term "neural tube defect" to designate anencephaly, encephalocele, and myelomeningocele. A third possibility is that a closed neural tube reopens, but supporting evidence is weakest in this area.
The most influential set of embryological standards, the Normal Plates of the Development of the Vertebrates, was edited in 1897 by the German zoologist, anatomist, and developmental biologist, Franz Keibel (1861-1929) (176). Keibel’s design adapted a plate by Swiss anatomist and embryologist Wilhelm His Sr (1831-1904) (145; 176; 153). As early as 1874, His compared the closure of the neural tube to a slit rubber tube.
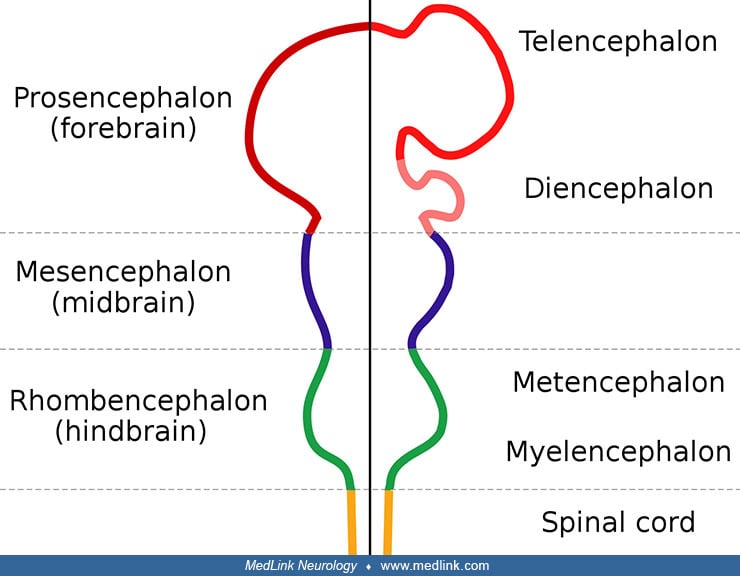
In human embryos, primary neurulation (closure of the neural folds) takes place between stage 8 and stage 12 (see Table 1). Anencephaly and other neural tube defects occur when development is abnormal, although the exact mechanism of closure remains unknown. The Hedgehog signaling pathway has been implicated in neural tube closure because increased Sonic Hedgehog signaling is associated with the appearance of exencephaly (230). Embryonic metabolism also appears to be important because studies using mass spectrometry have identified abnormalities in glucose metabolism during neural tube closure (379).
Some investigators have postulated that the variations of anencephaly and other neural tube defects can be explained by multiple sites of closure of the neural tube (352; 125; 325). Others counter that “accessory loci” for closure exist in human embryos but are highly variable and do not follow a constant pattern (254). Research progress is intermittent and capricious because affected embryos are rare and die at very early ages (ie, 5 to 6.5 weeks) (231). Two anencephalic embryos, stages 13 and 22, are extreme exceptions to this, having intact neural crest derivatives (255). This observation suggests that the process responsible for anencephaly began after the initiation of crest cell migration or that crest cells survived the pathogenetic event.
|
Stage |
Age* |
Pertinent event |
Results of maldevelopment |
|
8 |
18 days |
Neural folds appear |
Brain and spinal cord exposed and dysplastic (ie, early craniorachischisis) |
|
9 |
20 days |
Neural groove |
Early craniorachischisis |
|
10 |
22 days |
First fusion of neural folds |
Craniorachischisis |
|
11 |
24 days |
Closure of rostral neuropore optic vesicle |
Anencephaly |
|
12 |
26 days |
Closure of caudal neuropore; primary neurulation ends in this stage or stage 13 |
Myelomeningocele; myelocele; encephalocele |
|
| |||
An understanding of these stages is important for counseling parents. Anencephaly can be said to have a "termination point" (ie, a time after which the anomaly cannot occur); therefore, exposures or other maternal events that occur after the critical periods for anencephaly can be ruled out as causative factors.
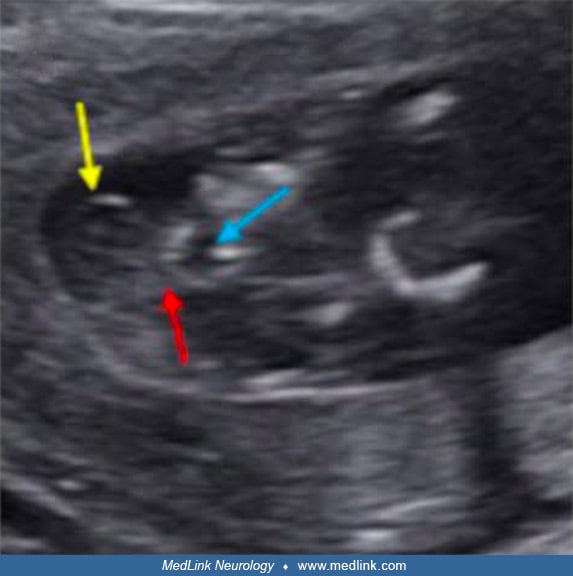
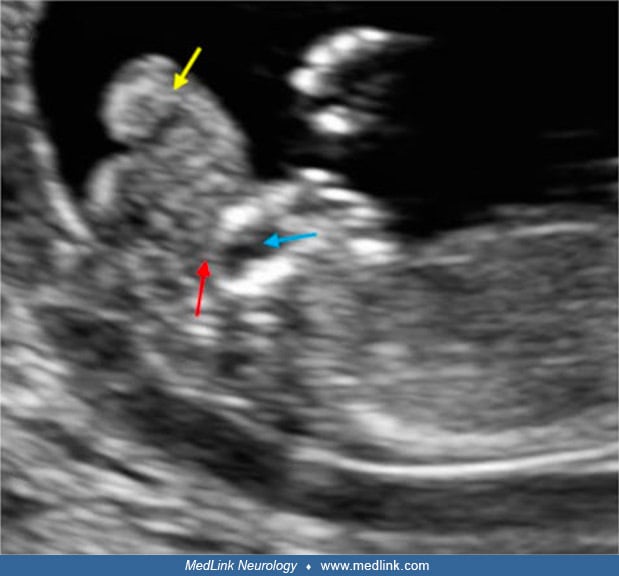
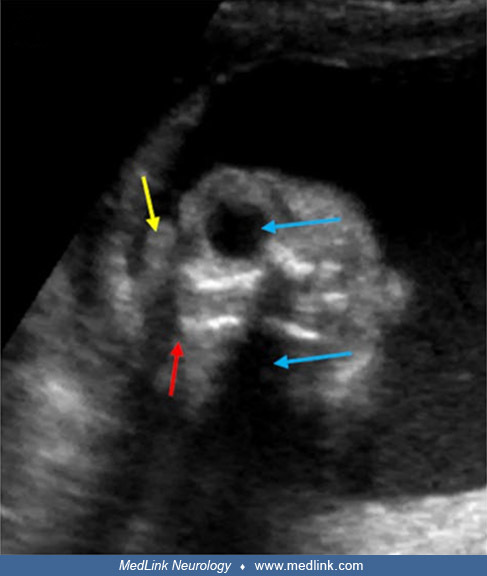
Exencephaly, the superior extrusion of a relatively large, bulbous portion of cerebral tissue beyond the calvaria, is a forerunner of anencephaly (376; 229). This has been demonstrated experimentally and by prenatal ultrasonography in fetuses before spontaneous degeneration of brain tissue. Holoprosencephaly is rarely associated with anencephaly (190; 317) and may involve aberrant morphogenesis of prechordal and paraxial developmental fields (318). The genetics of neural tube defects and midline craniofacial malformations are becoming understood, as illustrated by studies of the tuft mouse mutant (115).
Pathological findings in the central nervous system depend on the gestational age and extent of the lesion. Rudimentary cerebral hemispheres can be found in 50% of anencephalic newborns, and complete absence can be found in the remainder. Absence of the cerebellum and brainstem has also been reported (356). These infants usually manifest a connective tissue mass adherent to the dura mater. Their cerebral tissue is soft and infiltrated with blood, and scattered islands of neural cells and choroid plexus can often be identified. As a result of this appearance, the term "area cerebrovasculosa" is often applied to the anencephalic brain. This vascular finding is a reactive change to the exposure of neural tissue to amniotic fluid.
Examination of the brainstem reveals variability in the preservation of nuclei and absence of the pyramidal tract. The eyes often bulge dramatically. They may also appear normal but lack central connections to the brain, ending blindly posterior to the optic foramina. Other pathological changes of the eyes include absence or reduction in axis cylinders and colobomata of the optic disc. The spinal cord may show reduced white matter in some tracts, absence of Clarke column, and a lower termination of the conus medullaris.
Cranial bones are highly abnormal (112; 229): calvarial bones are displaced, rudimentary, or absent; the zygomatic bones may be rotated abnormally; the sphenoid is hypoplastic in the cranial base; and petrous ridges are malpositioned. The maxillae, palatine bones, and vomer may be normal, although the mandible is large and prognathic. Temporal and occipital bones may also be malformed (13). An understanding of this bony anatomy has proven especially helpful in certain forensic situations involving found human remains of anencephalic infants (97).
Additional malformations of other systems are common. One study found that liveborn anencephalics had 12% additional malformations of other systems, but if there was an associated spina bifida or encephalocele, the prevalence was 88% (294). Anomalies of extraocular muscles have been observed in a large percentage of affected fetuses (268). Endocrine system anomalies are present in anencephalic infants with absence or hypoplasia of the pituitary gland and nearly universal hypoplasia of the adrenal glands (212). Diaphragmatic malformations (eg, hernia or eventration), abdominal wall defects, and other thoracic cage abnormalities are common (189; 221). Frequently, the lungs and heart are hypoplastic. One study demonstrated various degrees of intestinal aganglionosis in anencephaly (209). Anthropometric studies have shown an increased growth of the arms in anencephaly (232). A proximodistal gradient existed, with the upper arm being increased by 24%, the forearm by 16%, and the hand by 2%. This is a variable finding, for Barr has observed normal arm length in anencephaly (Barr, personal communication 1997). Other anomalies of the limbs are found, but no one abnormality stands out, except perhaps talipes equinovarus. One detailed anatomic study identified several anomalies of skeletal muscles in a single fetus (06). A sternalis muscle is present in up to 50% of anencephalics but is unusual in the general population (1% to 4%).
Research directions. Important avenues for research include studies of genes controlling folate metabolism (87; 29), screens for human homologs to genes that cause anencephaly in laboratory animals (329), and maternal or sex-influenced genetic effects (84). Mutations in planar cell polarity genes (responsible for maintaining epithelial orientation during neural tube closure) have been identified in mice with neural tube defects and may be functional in humans as well (86; 167). The regulatory effects of bone morphogenetic protein and Sonic Hedgehog have been implicated in neural tube bending and may play a role in pathogenesis (67), as may maladjustment of microRNAs in the mitogen-activated protein kinase signaling pathway (390).
Of congenital defects, only cardiac defects are more common than neural tube defects. Over the past 50 years, the prevalence of anencephaly has averaged about 0.5 to 2 per 1000 deliveries, but wide variations exist geographically and by race (65; 170). A meta-analysis of 360 studies with a collective sample size of 207,639,132 people estimated the prevalence of anencephaly worldwide at 0.5 per 1000 births (289). Worldwide, the annual number of new cases is estimated to be at least 214,000 to 322,000 (208; 170).
In the United States, among 13 population-based programs from 2016 to 2020, the estimated prevalence per 10,000 live births was 2.28 for anencephaly, 1.04 for encephalocele, and 3.65 for spina bifida without anencephaly (note the change in units with the preceding paragraph as rates in the U.S. are below the world averages) (327). Prevalence of anencephaly, encephalocele, and spina bifida without anencephaly varied by race and ethnicity (327). Compared to non-Hispanic whites, rates of anencephaly were significantly higher among Hispanics in the U.S. The estimated prevalence per 10,000 live births was 1.75 for non-Hispanic whites, 1.77 for non-Hispanic blacks, 2.34 for Hispanics, and 1.90 for non-Hispanic Asians or Pacific Islanders. Compared to non-Hispanic whites, rates of encephalocele were significantly higher among non-Hispanic blacks in the U.S. The estimated prevalence of encephalocele per 10,000 live births was 0.87 for non-Hispanic whites, 1.54 for non-Hispanic blacks, 0.97 for Hispanics, and 0.45 for non-Hispanic Asians or Pacific Islanders. Finally, compared to non-Hispanic whites, rates of spina bifida without anencephaly were significantly lower among non-Hispanic blacks and non-Hispanic Asians or Pacific Islanders and significantly higher among Hispanics in the U.S. The estimated prevalence of spina bifida without anencephaly per 10,000 live births was 3.68 for non-Hispanic whites, 2.35 for non-Hispanic blacks, 4.21 for Hispanics, and 1.94 for non-Hispanic Asians or Pacific Islanders. Rates were also presented for non-Hispanic American Indian or Alaska Native population, but the population size and counts are quite low, making rates less reliable.
Reported neural tube defect birth prevalence figures do not include the large number of anencephalic embryos and fetuses that undergo spontaneous abortions and may not include therapeutic terminations (27; 107; 193; 265). In fact, reports of the declining birth prevalence of neural tube defects may be influenced significantly by pregnancy termination (337). An estimated 83% of pregnancies complicated by anencephaly and 63% of cases with spina bifida undergo termination (163). The termination rate in France is estimated at 97% (342). Data from these cases may not be available in the form of death certificates or other formal documentation. These points are borne out by a study in Japan, wherein an extensive review of over 311,000 deliveries and terminations in cases of myelomeningocele and anencephaly disclosed a birth prevalence of 0.8 to 0.9 per 1000 deliveries (2014 and 2015 data) (181). Pregnancies involving anencephaly were terminated in 80%, an indication of the potential magnitude of data that may be lost if only live births are reported.
Birth prevalence may also fluctuate over time and among different cultures. British Columbia, for example, has experienced a decline in the occurrence of neural tube defects (but also an increase in the severity of lesions) and a striking decrease in recurrence from 2.3% to 0.2% (56). From 1997 to 2019, the overall prevalence of neural tube defects in Canada was stable at 0.74 per 1000 total births (197). The birth prevalence in Ireland (where terminations were illegal) dropped 4-fold from 1980 (4.7 per 1000 births) to 1994 (1.2 per 1000 births) (215). This resembles the prevalence for Germany (1.2 per 1000 births), a country that has lagged in folate fortification (142). By contrast, a decrease in birth prevalence in northern England is attributed to improved prenatal diagnosis and intervention (274).
Some less developed countries with lower rates of periconceptional folic acid supplementation and without food fortification with folic acid have higher rates of neural tube defects. For example, in a cross-sectional study in northwestern Nicaragua from 2006 to 2018, the prevalence of neural tube defects was 14.0 per 10,000 births (95% CI: 12.3-15.7), with corresponding values for spina bifida at 7.8 per 10,000 births, anencephaly at 5.4 per 10,000 births, and encephalocele at 0.7 per 10,000 births (263). Mothers with fetuses or newborns with neural tube defects did not use folic acid before conception, and 11% experienced periods of hyperthermia during the first trimester of pregnancy.
Considerable variation has been demonstrated for neural tube defects in Africa (326).
(Source: Ssentongo P, Heilbrunn ES, Ssentongo AE, Ssenyonga LVN, Lekoubou A. Birth prevalence of neural tube defects in eastern Africa: a systematic review and meta-analysis. BMC Neurol 2022;22[1]:202. Creative Commons Attribut...
(Source: Ssentongo P, Heilbrunn ES, Ssentongo AE, Ssenyonga LVN, Lekoubou A. Birth prevalence of neural tube defects in eastern Africa: a systematic review and meta-analysis. BMC Neurol 2022;22[1]:202. Creative Commons Attribut...
(Source: Ssentongo P, Heilbrunn ES, Ssentongo AE, Ssenyonga LVN, Lekoubou A. Birth prevalence of neural tube defects in eastern Africa: a systematic review and meta-analysis. BMC Neurol 2022;22[1]:202. Creative Commons Attribut...
(Source: Ssentongo P, Heilbrunn ES, Ssentongo AE, Ssenyonga LVN, Lekoubou A. Birth prevalence of neural tube defects in Eastern Africa: a systematic review and meta-analysis. BMC Neurol 2022;22[1]:202. Creative Commons Attribut...
(Source: Ssentongo P, Heilbrunn ES, Ssentongo AE, Ssenyonga LVN, Lekoubou A. Birth prevalence of neural tube defects in eastern Africa: a systematic review and meta-analysis. BMC Neurol 2022;22[1]:202. Creative Commons Attribut...
An increased birth prevalence of anencephaly has been noted among Hispanics in South Texas. Hispanics have a birth prevalence three times higher than non-Hispanics (37), though no difference exists between Hispanics born in Mexico and Hispanics born in Texas (38). This difference in birth prevalence has also been noted in California, where Hispanic women were 45% more likely to have an anencephalic conceptus than white women (111) and elsewhere in the United States (26).
Although international and national distributions of neural tube defect occurrence have often been used to generate etiological hypotheses concerning neural tube defects (eg, potato blight, tea consumption, and zinc deficiency), few of these studies have been conducted with scientific rigor, and most have imprecise estimates due to small sample sizes (24). Moreover, comparability is compromised by nonuniformity in the duration and diligence of case ascertainment, the lack of a standardized nomenclature and classification, inconsistency in the placement of the gestational boundary between late fetal deaths and spontaneous abortions, and differences in the choice of the denominator (24). Findings are often compared from studies conducted at different times without considering secular trends and using studies with varying thoroughness of case ascertainment, all inherently assuming that the differences reflect true spatial or temporal variations rather than variations in the quality of ascertainment. Consequently, geographic patterns of neural tube defects may be attributable to variations in the validity of studies used to describe these patterns (24).
The rate was four times the national average in a central Washington state cluster of neural tube defects, mostly anencephaly (51). In this study, no significant differences were found between cases and controls. The area is largely rural, so an effect of pesticides and other agricultural materials might be postulated. However, an epidemiological study from California did not identify an association between pesticide exposure and neural tube defects (381).
Death certificate reports of anencephaly may be imprecise or inaccurate (312). The diagnosis of anencephaly on death certificates relies on the expertise and experience of the individual completing the document. Fetuses dying by natural means or pregnancy termination before 20 weeks elicit no death certificate. By one estimate, over one third of anencephalic fetuses may not be included in published frequencies, which, for this reason, are low (366).
The etiology of neural tube defects is complex, with both genetic (polygenic or oligogenic) and environmental factors having important contributions (66).
Dietary folate. Dietary folate intake is the strongest known and best-documented risk factor for neural tube defects (184).
Observational studies. The prevalence of anencephaly has increased during periods of poor maternal nutrition (eg, the Great Depression and wartime famines), and it has dropped with global socioeconomic development (225).
In 1964, British obstetrician Brian Hibbard (1926-2021) suggested an association between fetal neural tube defects and maternal deficiency or defective metabolism of folates (144; 184). In 1976, Richard Smithells (1924-2002) and colleagues at the University of Leeds demonstrated that women with megaloblastic anemia during pregnancy have a high frequency of neural tube defects in their offspring (321). In 1980, Smithells and colleagues reported a nonrandomized trial of multivitamin supplementation among women who had previously given birth to one or more infants affected with neural tube defects (322); there was a 5% recurrence rate for the nonsupplemented group compared with a 0.6% recurrence rate for the supplemented group.
Additional observational studies and nonrandomized clinical trials were published during the 1980s and 1990s and documented the protective effects of higher folic acid intake or of vitamin supplements containing folic acid during the periconceptional period (ie, from 1 month before pregnancy through the first trimester) among women who had not previously had a pregnancy affected by a neural tube defect (ie, occurrence studies) and among women who had a previous pregnancy affected by a neural tube defect (ie, recurrence studies) (184). These studies showed a wide range of estimated efficacy in the occurrence of neural tube defects with folic acid supplementation, but the summary efficacy estimate across the various studies indicated an overall 50% reduction in risk of neural tube defects (360).
Using fetal death records in Argentina between 1994 and 2019, fetal deaths due to anencephaly and myelomeningocele decreased significantly between 2005 and 2019 (by 67% and 51%, respectively) compared with the period between 1994 and 2004 before the effective fortification of wheat flour (31). In 1994, neural tube defects accounted for 34.7% of congenital malformation fetal deaths and 1.7% of all fetal deaths, whereas in 2019, these percentages were 9.4% and 0.5%, respectively.
Randomized controlled trials. The strongest evidence in support of periconceptional folic acid supplementation comes from two large, randomized trials published in the early 1990s (227; 77; 72; 74; 73; 360; 79; 184). The Medical Research Council (MRC) study under the direction of Sir Nicholas Wald (b 1944) at St. Bartholomew’s Hospital Medical College in London was a multicenter, multinational, randomized, double-blind, controlled, recurrence prevention trial conducted in 33 centers in seven countries (227; 360). The MRC study found a 72% reduction in recurrence of neural tube defects with 4 mg of folic acid daily from before conception and during the first trimester among women with a previous neural tube defect-associated pregnancy (227; 360). The Hungarian study, conducted by Andrew Czeizel (1935-2015) of the National Institute of Hygiene in Budapest, was a randomized, double-blind, controlled occurrence prevention study with periconceptional supplementation with multivitamins, including 0.8 mg of folic acid (77; 79; 73; 362). Among approximately 5000 women with confirmed pregnancy and an “informative offspring,” maternal periconceptional folic acid supplementation significantly decreased the first occurrence of neural tube defects compared to a placebo-like (ie, trace element) control group.
A meta-analysis of data from these trials and a previous small (underpowered and not statistically significant) trial by Laurence and colleagues collectively indicated that periconceptual folate administration reduces both the occurrence and recurrence risks of neural tube defects by at least 70% (185; 227; 360). Subsequent studies have generally supported these findings and suggest that periconceptional multivitamin supplementation can significantly reduce the occurrence of other congenital abnormalities in addition to neural tube defects (179; 72; 74; 78).
Governmental recommendations and health policy. Data from the randomized controlled trials have been used to establish (1) governmental recommendations concerning folic acid intake and (2) health policy concerning vitamin fortification of foodstuffs (148; 184). In 1991, the U.S. Centers for Disease Control and Prevention (CDC) published a review of the evidence for the prevention of recurrent neural tube defects and recommended 4 mg of folic acid for women who had previously had an infant or fetus with a neural tube defect (50). In 1992, the U.S. Public Health Service recommended that all women capable of becoming pregnant should consume 0.4 mg (4000 mg) of folic acid daily (47; 69). Because naturally occurring folate is less readily absorbed than synthetic folic acid, in 1998, the Institute of Medicine recommended that women of childbearing age consume 0.4 mg daily from dietary supplements or fortified foods for the primary prevention of neural tube defects (156).
Potential strategies for increasing folate levels among women are (1) dietary modification, (2) folic acid supplementation, and (3) food or salt fortification (47; 363; 75; 217; 184; 264). Despite various education campaigns, the estimated dietary folate intake for U.S. women averages only 0.2 mg daily, and it was considered impractical to have women systematically increase their intake of folate-rich foods (eg, fruits, leafy green vegetables, and grains) sufficiently to raise daily folate intake to 0.4 mg daily (46). Folic acid supplementation can also be effective, but vitamins are consistently used by less than a third of women of childbearing age, and the remainder do not consider taking vitamin supplements until after they discover that they are pregnant (217; 353).
(Source: van der Windt M, Schoenmakers S, van Rijn B, Galjaard S, Steegers-Theunissen R, van Rossem L. Epidemiology and (patho)physiology of folic acid supplement use in obese women before and during pregnancy. Nutrients 2021;1...
Unfortunately, neural tube defects develop in the fourth week after conception, when most women are unaware of their pregnancies. Furthermore, about half of the pregnancies are unplanned (129; 121), but even women who plan their pregnancies are poorly compliant with folate supplementation (63; 299; 370; 217). Therefore, an approach relying on supplementation will not prevent most of the folate-preventable cases of neural tube defects. Food fortification, in contrast, can cost-effectively increase folate levels across the population without requiring a change in behavior (281; 362).
Food fortification. In 1996, the U.S. Food and Drug Administration (FDA) selected flour, corn meal, pasta, and rice for mandatory folic acid fortification beginning in January 1998 at a level of 140 mg per 100 g of cereal grain product (118; 120; 117; 119; 184). This was estimated to result in an average adult consumption of approximately 100 mg of folic acid daily from fortified cereal grain products, effectively ensuring that about half of women of reproductive age would receive the recommended 0.4 mg daily from all sources (118; 120; 281). This level of fortification--considered the best possible accommodation between concerns for the fortification of the target population of women of childbearing age and the safety of the much larger no-target population--was expected to prevent many but not all neural tube defects that might be prevented by sufficient maternal folic acid intake. Folic acid fortification was limited to relatively low levels because of a fear that folic acid would correct the hematological abnormality in patients with vitamin B12 deficiency, potentially delaying diagnosis and allowing progression of central and peripheral nervous system manifestations of vitamin B12 deficiency (279). Many have challenged the logic and ethics of this rationale and the resulting national fortification decisions (364), but fortification levels remain modest. As a result, dietary modification and folic acid supplementation continue to be necessary and appropriate modes of intervention.
Since 1996, folic acid fortification has produced a significant improvement in population folate status in the United States (160; 186; 148; 362; 103; 210; 272; 48; 266; 184).
Prevalence of neural tube defects (anencephaly and spina bifida) before and after mandatory folic acid fortification, by maternal race/ethnicity--19 population-based birth defect surveillance programs, United States, 1995–2011....
Neural tube defect rates per 10,000 population, by race/ethnicity and fortification period status. Data from 25 population-based birth defect programs. In 1996, Food and Drug Administration established regulations requiring for...
In 1999, data from the Framingham Offspring Cohort showed that fortification of enriched grain products with folic acid was associated with a substantial improvement in the folate status of the population (160; 272). Similar results were demonstrated in populations enrolled in large managed care plans (186) and in representative samples of women participating in the National Health and Nutrition Examination Survey (NHANES) (44; 266).
By 2001, findings using birth certificate data for live births in 45 states and the District of Columbia between 1990 and 1999 suggested that a decline of approximately 20% in the prevalence of neural tube defects at birth followed fortification of the U.S. food supply with folic acid (148). A later analysis by the CDC suggested a 27% decline in the average annual proportion of pregnancies affected by neural tube defects after fortification (ie, 1999-2000 compared with 1995-1996) (48).
Folic acid supplementation has been successful in reducing the prevalence of defects in Hispanic and non-Hispanic white patients but not necessarily in black patients (219; 331; 122; 372). The fortification of corn masa flour, a food used in Hispanic food preparation, could be useful in further reducing neural tube defects in Hispanics (131; 228).
Similar improvements have been observed in other countries that have adopted this strategy (276; 362; 42; 88; 295; 280). For example, a 31% decrease has been noted in Chile (42). In Brazil, there has been a 30% decrease in the prevalence of neural tube defects and a 23% decrease in hospital and outpatient costs associated with caring for people with neural tube defects following mandatory flour supplementation (295; 280). In Canada, the prevalence of neural tube defects decreased by 46% from 1.58 per 1000 births before fortification to 0.86 per 1000 births during the full fortification period (88).
Overall, fortification of flour with folic acid has proven to be an effective approach for preventing pregnancies with neural tube defects and a cost-effective strategy for decreasing overall costs of care for neural tube defects (280). Unfortunately, only 69 countries have mandatory fortification of grains that includes folic acid (359). Among countries with adequate data available, 63,520 cases of spina bifida and anencephaly were prevented in the year 2022 in 68 countries implementing mandatory folic acid fortification of grains with folic acid (359). Because of the incomplete adoption of food fortification, less than a quarter (23.7%) of all possible folic acid-preventable spina bifida and anencephaly cases are, in fact, prevented globally (359).
Folic acid is also successful in preventing recurrent neural tube defects (361; 275; 331).
Mechanism of folate action in preventing neural tube defects. The physiological effects of folic acid on embryological development are complex and incompletely understood. There is general agreement among several observational studies that folate deficiency, hyperhomocysteinaemia, and homozygosity for the methylenetetrahydrofolate reductase thermolabile variant are probable risk factors for placenta-mediated diseases, such as preeclampsia, spontaneous abortion, and placental abruption (277).
For example, an association between deficiencies in folate level and increased pregnancy loss has been suggested (277), but agreement on this issue is not universal (335). The idea that folic acid supplementation reduces the prevalence of neural tube defects by inducing miscarriage in affected fetuses has been proposed (149; 150; 149; 76) and challenged (32; 130; 298; 277; 21). Available data in humans have not resolved the debate, but studies using mouse models do not support the proposed action of folic acid in encouraging the in-utero demise of affected fetuses (ie, "terathanasia") (66). Incidentally, the word "terathanasia" has been introduced and promulgated but is a misnomer; the correct term should be “teratothanasia” (124).
Antifolates: substances that interfere with folate metabolism. A diet rich in folate may lower the risk of neural tube defects, whereas conversely, food substances, toxins, and pharmaceuticals that interfere with folate metabolism may increase risk (194; 40).
Tea catechins. Compounds called catechins in tea have antifolate properties and, for this reason, have been evaluated as potential risk factors for neural tube defects (237; 236; 239; 238; 241; 05; 292; 293; 291; 324; 14; 315; 288; 287; 378; 385; 384; 330; 391; 350; 380).
Flavonoids present in tea with epicatechin (ECG, EGCG) and catechin configurations (CG). Catechins are major components of tea. Catechins contain a benzopyran skeleton with a phenyl group substituted at the 2-position and a hyd...
Structural comparison of natural (ECG) and synthetic (TMECG) polyphenols with classical (MTX) and non-classical (TQD) antifolates. Abbreviations: ECG: (-)-epicatechin gallate; MTX: methotrexate; TMECG: 3-O-(3,4,5-trimethoxybenz...
Wall-eyed stereo view of ECG modeled into the folate-binding site of human dihydrofolate reductase (DHFR). Abbreviations: ECG: (-)-epicatechin gallate; DHFR: dihydrofolate reductase. Carbon atoms of the ECG ligand and surroundi...
The ester bond containing tea polyphenols epigallocatechin gallate (EGCG) and epicatechin gallate (ECG) are potent and specific inhibitors of dihydrofolate reductase (DHFR) activity in vitro at concentrations found in the serum and tissues of green tea drinkers (239; 240; 293; 288) and can also downregulate DHFR expression (291). Their slow-binding inhibition of DHFR is thought to be due to the formation of a slow dissociation ternary complex by the reaction of nicotinamide adenine dinucleotide phosphate (NADPH) with the enzyme-inhibitor complex (239; 240). Catechins contained in green tea also apparently inhibit the cellular uptake of folic acid (05).
The anti-inflammatory and anti-cancer properties of EGCG are mediated by folate cycle disruption, adenosine release, and NF-kappaB suppression (239; 240; 2007; 2008; 292; 378). EGCG inhibits the growth of a human colon carcinoma cell line in a concentration- and time-dependent manner (239; 240). Rescue experiments using leucovorin and hypoxanthine-thymine medium indicated that EGCG can disturb folate metabolism within cells (239). In addition, EGCG increased the uptake of tritiated thymidine and showed synergy with 5-fluorouracil, whereas its inhibitory action was strengthened after treatment with hypoxanthine, which indicates that EGCG decreases the cellular production of nucleotides, disturbing both DNA and RNA synthesis (239; 240). In addition to its effects on nucleotide biosynthesis, it is linked to a decrease in cellular methylation as a result of folic acid deprivation, and it causes adenosine to be released from the cells because it disrupts purine metabolism (239; 240).
Antiviral activities of EGCG with different modes of action have been demonstrated on diverse families of viruses, including interfering with the replication cycle of DNA viruses (330). Most of these in vitro studies demonstrated antiviral properties within the range of physiological concentrations of EGCG. In contrast, the minimum inhibitory concentrations against bacteria are generally 10- to 100-fold higher than those needed to demonstrate antiviral properties (330).
Nevertheless, EGCG affects the folic acid metabolism of bacteria and fungi by inhibiting the cytoplasmic enzyme DHFR (237; 324; 330). EGCG acts as a bisubstrate inhibitor on bacterial DHFR, with the ability of EGCG to bind to the enzyme both on substrate (DHF) and cofactor (NADPH) sites (324). The catechin EGCG, one of the main constituents of green tea, showed strong antibiotic activity that was attributable to its antifolate activity, which was synergistic with the nonclassical antifolate compound, trimethoprim, and attenuated by including leucovorin in the growth medium (237). Moreover, the tea polyphenol epigallocatechin-3-gallate inhibits ergosterol synthesis by disturbing folic acid metabolism in Candida albicans (236).
Some studies have suggested that higher tea intake during the periconceptional period was associated with an increased risk of neural tube defects (385; 384), but data at present are inconsistent and insufficient to either strongly support or exclude this possibility.
In a human pilot study, green tea extracts lowered serum folates in rats at very high dietary concentrations but did not affect plasma folates (14).
In a health study of pregnant women with singleton pregnancies in Japan, the serum folate levels of the participants with high consumption of green tea or oolong tea were significantly lower than those of others after adjusting for confounding variables, including dietary folate intake and use of folic acid supplements or multivitamins (315).
In a study from China utilizing a population-based surveillance system in northern China from 2002 to 2007, 631 neural tube defect cases and 857 controls were included (385). Compared with women who did not drink tea during the periconceptional period, women who drank tea daily had a 3-fold increased risk of having a neural tube defect-affected pregnancy (odds ratio = 3.1; 95% CI = 1.4-7.0). The elevated risk associated with daily tea drinking remained after adjusting for maternal age, educational level, occupation, and periconceptional folic acid supplementation. The association was present for major subtypes of neural tube defects (ie, anencephaly, encephalocele, and spina bifida).
In a subsequent study using data collected in the Slone Epidemiology Center Birth Defects Study from 1976 to 2010, there was no significant overall increased risk for daily tea intake. However, for 1998 and onward, there was a suggestion of a modest increase for those who drank more than three cups a day, although this did not reach statistical significance (OR 1.92; 95% CI 0.84-4.38). Among women with total folic acid intake greater than 400 μg, consumption of three cups or more of tea per day was associated with an increased risk of spina bifida during the period 1976 to 1988 (OR 2.04; 95% CI 0.69-7.66) and in later periods (OR, 3.13; 95% CI, 0.87-11.33), but none of these were statistically significant.
A systematic review and meta-analysis found that tea consumption in the periconceptional period does not significantly increase the prevalence of neural tube defects (380). Seven articles with nine studies collectively comprised 2834 cases and 19,924 participants. Tea consumption during the periconceptional period did not significantly increase neural tube defect prevalence (OR 1.37; 95% CI 0.96-1.95), a finding that was consistent for three subtypes of neural tube defects: anencephaly (OR 1.36; 95% CI 0.84-2.20), spina bifida (OR 1.51; 95% CI 0.84-2.72), and encephalocele (OR 0.99; 95% CI 0.46-2.15). Furthermore, no dose-response association between tea consumption and the risk of neural tube defects was evident.
Pharmaceuticals. Exposure to folic acid antagonists or "antifolates" (ie, carbamazepine, phenobarbital, phenytoin, primidone, sulfasalazine, triamterene, and trimethoprim, methotrexate, aminopterin) increases the risk for neural tube defects (141). Women taking antiepileptic drugs are at an increased risk of having offspring with neural tube defects, which is at least partly due to the antifolate properties of many anticonvulsants (113). Folic acid supplementation does reduce, but does not eliminate, the risk of neural tube defects in women taking these drugs (122).
Folate-receptor autoantibodies. A small case-controlled study of women from Brooklyn, New York, found that serum from women with a pregnancy complicated by a neural tube defect contained autoantibodies that bind to folate receptors that can block the cellular uptake of folate (283). However, the presence and titer of maternal folate-receptor autoantibodies in stored frozen blood samples were not significantly associated with a neural tube defect-affected pregnancy in an Irish population (226). A separate, smaller, case-controlled study using fresh samples produced similar results. A subsequent population-based cohort study utilizing samples obtained from the California Birth Defects Monitoring Program found that high concentrations of IgG or IgM antibodies to folate receptors or folate binding proteins in midgestational serum samples were significantly associated with neural tube defects (33). Therefore, high concentrations of IgG or IgM antibodies to folate receptors or folate-binding proteins are risk factors for neural tube defects.
Folate-resistant neural tube defects. Recurrence of neural tube defects despite high-dose folic acid supplementation suggests that a proportion of neural tube defect cases are "resistant" to folic acid, or at least are not amenable to folate supplementation, and presumably occur by other (possibly genetic) mechanisms (43). Heterogeneity in the etiology of neural tube defects has also been suggested in animal studies, particularly mouse models.
Inositol isomers. Inositol metabolism is associated with neural tube defects, but the mechanisms are not clear (138; 137; 139; 355; 66; 43; 393; 95; 158).
Inositol can be obtained from dietary sources or synthesized from D-glucose by the sequential action of hexokinase (HK), myo-inositol-1-phosphate synthase (MIPS), and inositol monophosphatase (IMPas). The structures of myo-inos...
Inositol isomers (particularly myo- and chiro-inositol) can prevent folate-resistant neural tube defects in the curly-tail mutant mouse model, suggesting that some cases of human neural tube defects might also benefit from inositol supplementation (66; 43; 95). Specific isoforms of protein kinase C are essential for the prevention of folate-resistant neural tube defects by inositol; specifically, there is an absolute dependence on the activity of PKCbeta and gamma for the prevention of neural tube defects by inositol and partial dependence on PKCzeta (64).
Pregnant women carrying fetuses with neural tube defects have lower concentrations of inositol in their blood than pregnant women not carrying fetuses with neural tube defects (43). Moreover, in pilot studies, periconceptional combination therapy with folic acid and inositol produced normal live births despite a high recurrence risk for neural tube defects (43). Inositol may be used prophylactically in humans in conjunction with folate supplementation (68; 344).
Valproic acid embryopathy. Valproic acid has been linked to neural tube defects in humans and in various animal models (346; 234; 250; 132; 297; 258; 04; 71; 259; 323; 252; 83; 85; 260; 161; 348; 347; 104; 200; 341; 03; 154; 349; 308; 309; 307; 267; 328). In humans, valproic acid is associated with a 1% to 2% birth prevalence of neural tube defects when taken during the first trimester of pregnancy. A high frequency of neural tube defects following maternal exposure has been demonstrated in mammals (particularly mice) as well as in birds and amphibians (250; 349).
Mouse models. In mouse models, maternal valproic acid treatment reduced the number of live fetuses per litter, impaired fetal growth, and increased the frequency of major malformations, including, most prominently, exencephaly (04).
Maternal valproic acid treatment produced gross metabolic changes in embryos, likely caused by a multiplicity of mechanisms, including the following:
|
• Interference with folate metabolism (346; 100; 101; 234; 146; 259; 323; 83) through methionine deficiency inhibition and interference with homocysteine catabolism (146) | |
|
• Markedly perturbed purine and pyrimidine metabolism (03) | |
|
• Disruption of mitochondrial beta-oxidation (03) | |
|
• Deregulation of apoptosis during development (348; 347; 200) | |
|
• Altered metabolism of sulfur-containing amino acids and glutathione (146) | |
|
• Diminished supra-cellular F-actin cables, which normally run along the lateral neural folds, and prevention of caudal narrowing of the posterior neuropore, in effect preventing morphological and biomechanical conversion of the caudal extreme of the posterior neuropore during late spinal closure (154) | |
|
• Cell cycle inhibition and alterations of mitotic rate (205; 348) | |
|
• Alterations of intercellular adhesion (205) | |
|
• Redox disruption (267) | |
|
• DNA damage and deregulation of multiple genes (348; 347) |
By comparing the effects of maternal exposure to valproic acid and various enantiomers, structure-activity relationships show a strict structural requirement for high teratogenic potency: the molecule must contain an alpha-hydrogen atom, a carboxyl function, branching on carbon atom 2 with two chains containing three carbon atoms each for maximum teratogenic activity (235). Because pharmacokinetic studies demonstrated that the various enantiomers reached the embryo to the same degree, the intrinsic teratogenic activity of the enantiomers differ. Therefore, stereoselective interaction between valproic acid and a chiral structure within the embryo is involved in the mechanism of teratogenicity. In contrast, the anticonvulsant activity of valproic acid and related compounds does not have this property, offering the possibility to develop novel antiepileptic agents with low teratogenic potency.
The peak concentrations of valproic acid, not the total dose provided (ie, as indicated by the area under the concentration/time curves), correlated with the teratogenicity of this compound (233).
The valproic acid embryopathy was more common in some mouse strains, indicating that genetic factors moderate the effect of valproic acid (258; 71; 252). In particular, a gene for ribonucleotide reductase subunit R1 conferred sensitivity to valproic acid-induced neural tube defects in mice that specifically targets and disrupts neural tube closure between the prosencephalon and mesencephalon region (future fore/midbrain; neural tube closure site II) (71). Valproic acid also deregulates genes involved in the cell cycle and apoptosis pathways of neural tube cells (252; 347) and upregulates various transcription factors, including Stat3 (signal transducer and activator of transcription 3), a transcription factor that is activated via tyrosine phosphorylation, as well as other transcription factors associated with cell survival and anti-apoptotic mechanisms (308; 309; 307).
Pretreatment with some additional chemicals was demonstrated to be protective against valproic acid-induced neural tube defects in embryos in some mouse models, including folic acid (83), folinic acid (346), vitamin E (04), pantothenic acid (297; 83), and sildenafil (which prolongs nitric oxide signaling) (341). Folic acid and pantothenic acid protect mouse embryos from valproic acid-induced neural tube defects by independent, but not mutually exclusive, mechanisms, both of which may be mediated by the prevention of valproic acid-induced alterations in specific proteins involved in neurulation (83). In addition, spirulina (a biomass of cyanobacteria with purported strong antioxidant effects) is reported to be protective against valproic acid-induced neural tube defects in mice.
In other mouse or chicken models, folate derivatives were not protective against valproic acid-induced neural tube defects (132; 349). Whether this reflects differences in genetic susceptibility alone or a precise sensitivity window is unclear. Strain-dependent susceptibility has been repeatedly demonstrated (132; 259; 323; 328). In another mouse model, prolonged dosing of folic acid was associated with sustained elevation of plasma levels (ie, above control levels) and acceleration of neural tube closure (259). Maintenance of elevated levels was necessary to provide protection against valproic acid-induced neural tube defect development, suggesting that plasma folic acid and B12 must be maintained at high levels throughout organogenesis to protect embryos against valproic acid-induced neural tube defects. Enhanced susceptibility of folate-binding protein-2 (Folbp2) knockout mice to in-utero valproic acid exposure was demonstrated for only some dietary folate regimens, indicating "a relatively frail relationship" between that genotype and valproic acid-induced neural tube defects (323).
Other drugs and chemical substances may act synergistically to increase the frequency of neural tube defects in offspring of mice exposed to valproic acid, including methotrexate (100), trimethoprim (101), ethanol (99), and homocysteine (260).
Cell culture models. In cell culture models, exposure to valproic acid significantly increased homologous recombination events and intracellular reactive oxygen species (ROS) levels, and the latter were attenuated by preincubation with polyethylene glycol-conjugated (PEG)-catalase (85). Valproic acid may produce detrimental DNA damage through ROS-mediated double-strand breaks in DNA (85; 347). Although these can be repaired through homologous recombination, the process is error-prone; consequently, detrimental genetic changes may occur. Because the developing embryo requires tight regulation of gene expression to develop properly, loss or dysfunction of genes involved in embryonic development through aberrant homologous recombination may ultimately cause neural tube defects.
The valproic acid-induced gene expression response in cultured cells indicates that approximately 30% of the approximately 200 genes known from genetic mouse models to be associated with neural tube defects were altered in embryos of mothers exposed to valproic acid (161). Altered gene expression was demonstrated for other known effects of valproic acid (ie, histone deacetylase inhibition, G[1]-phase cell cycle arrest, induction of apoptosis). Therefore, combined deregulation of multiple genes is a possible mechanism of valproic acid teratogenicity.
Other risk factors. Overarching reviews of the epidemiology of neural tube defects suggest that a host of interactions, including gene-gene and gene-environment, as well as maternal genetic effects, probably affect the risk of neural tube defects (223; 282).
Obesity and gestational diabetes. In a study conducted in the United States, women at highest risk were young, unmarried, obese smokers who ate few fruits and vegetables and had a low level of education (45). Obesity and gestational diabetes remain risk factors even when adjustments are made for the effects of maternal age, education, ethnicity, vitamin use, and tobacco and alcohol use (09). A lack of periconceptional folic acid is associated with an increased risk for birth defects in women with diabetes mellitus (70).
Women who have undergone gastric bypass are at special risk, perhaps because obesity is a risk factor but also because folates and other vitamins are absorbed less completely following this procedure (224). Clearly, ongoing surveillance is necessary (19).
Parental age. An inverse association was observed between the age of the pregnant women and the occurrence of anencephaly in the Arkhangelskaja Oblast in Russia and in Norway (265). A meta-analysis of neural tube defects and maternal age found that advanced maternal age (ie, greater than 40 years) is also associated with an increased risk for spina bifida and, to a lesser extent, anencephaly, whereas mothers 19 years old or younger have a higher risk of having a child with spina bifida (357).
Paternal age does not appear to be a factor (12).
Occupational and environmental exposures. Further efforts are needed to assess various occupational and environmental exposures, particularly organic solvents, pesticides, and other chemicals used in agriculture, nitrates, heavy metals (eg, mercury), ionizing radiation, byproducts of water purification, and hazardous waste (306; 257; 183; 285). One paper found that paternal exposure to organic solvents is associated with an increased risk for neural tube defects (195). Another study found a positive association between maternal exposure to chlorinated solvents in early pregnancy and neural tube defects (90).
Maternal stress. Maternal stress, arising from challenging life events, has been associated with a 2.4-fold increase in risk for women who did not take folate supplements and a 1.4-fold increase in women who did (39).
Although the birth prevalence of anencephaly averages about 1 in 2000 live births, the recurrence risk is 100 times higher (about 1 in 20). Recurrence is higher for syndromic forms than isolated ones (60; 61). Many preventive efforts, therefore, have been directed toward pregnant women who have had a previous child with anencephaly or other neural tube defect.
Socioeconomic status. In the Texas-Mexico border region, an area recognized for an elevated incidence of neural tube defects, women living in poverty are at increased risk for having a baby with craniorachischisis (164).
Women employed in industry or agriculture have a 6.5-fold higher risk of having an anencephalic baby than business or professional women (23). A study identified no association between maternal alcohol consumption and anencephaly (196).
Residing in a census tract with more socioeconomic deprivation is associated with an increased risk for neural tube defects, specifically spina bifida (270).
Other potential risk factors. Other risk factors, including low serum B12, high serum homocysteine, diarrhea, stress, fumonisins (environmental toxins produced by Fusarium molds that grow on agricultural commodities in the field or during storage), high nitrate or nitrite intake, and obesity, play a role in the risk for Hispanics living on the Texas-Mexico border (333).
A 2018 study demonstrated an association between a low-carbohydrate diet and anencephaly or spina bifida (91); women with a carbohydrate-restricted diet were 30% more likely to have a baby with one of these defects. A follow-up study confirmed this association, indicating that the increased risk was not due to low folic acid intake (314).
Excess homocysteine may play an independent role in the development of neural tube defects (110; 333).
Neural tube defects are amenable to different prevention approaches. Anencephaly, specifically, is amenable to both primary and secondary prevention, whereas less severe forms of neural tube defects are also amenable to tertiary prevention.
Primary prevention. Primary prevention, which aims to prevent neural tube defects before they ever occur for a given woman, can be addressed by increasing periconceptional folate levels through (1) dietary modification, (2) folic acid supplementation, and (3) food fortification. Food fortification has been proven to be the most important and effective approach as it requires no change in behaviors. Moreover, because many women become aware of their pregnancies after the critical period has already begun, food fortification provides a safer background level of folate levels without requiring any active intervention or behavior change. Efforts are underway to monitor the use and efficacy of natural and exogenous forms of folic acid, with the goal of making both available worldwide (247; 248).
The use of both supplemental vitamins and fortified foods (ie, cereals) could contribute significantly to the reduction in neural tube defects (244; 245; 30; 148; 109; 206; 210; 374). However, physicians do not always understand the timing or dosage of folate administration (01).
Several trials worldwide have demonstrated the benefit of folic acid supplementation before and during early pregnancy in reducing the prevalence of first-time neural tube defects by as much as 70%. All women contemplating pregnancy should supplement their diet with 0.4 mg folic acid daily (364; 114), but, at present, only 15% to 25% of folic acid-preventable neural tube defects are being prevented globally (388). Increased intake of additional micronutrients, including betaine (trimethyl glycine), iron, and various vitamins [ie, A (including retinol), B1 (thiamine), B2 (riboflavin), B3 (niacin), B6 (pyridoxine), C, and E], may also decrease the risk of neural tube defects (57). Women aged 18 to 24 (of all educational levels) are least aware of the need for supplementation (52; 273).
Fortification of foodstuffs alone is insufficient as many women do not maintain protective folic acid levels, a situation further compounded by pregnancy, especially if there is hyperemesis gravidarum; its effects can, for example, be impacted adversely by dieting (53). The need for fortification is ongoing as women seem inconsistent in taking folic acid supplements during their childbearing years (49; 180), and making recommendations alone appears to be insufficient for prevention (332). Governing bodies are sometimes slow to develop policies as well (246).
Global prevention efforts for folic acid-preventable spina bifida and anencephaly are inadequate even after three decades of knowledge on their prevention (54; 172). Although mandatory food fortification with folic acid is a safe, cost-effective, and sustainable intervention to prevent spina bifida and anencephaly, only a quarter of all preventable spina bifida and anencephaly cases worldwide are avoided by food fortification (171). Universal mandatory fortification of staples should be urgently implemented to prevent thousands of folic acid-preventable spina bifida and anencephaly cases as well as associated elective terminations, stillbirths, and child mortality (172).
Harmful effects of folic acid supplementation have not been identified (358).
Secondary prevention. Secondary prevention, which aims to reduce the impact of neural tube defects that have already occurred, is accomplished by (1) detecting neural tube defects as soon as possible with prenatal screening and (2) encouraging personal strategies to prevent recurrence with subsequent pregnancies (eg, folate supplementation). Maternal serum and sonographic screening programs can identify most affected pregnancies, allowing parents to make decisions about pregnancy management.
Women who have a neural tube defect or a child with a neural tube defect should take 5 mg of folic acid daily (275).
Tertiary prevention. Tertiary prevention, which aims to lessen the impact of an ongoing illness or injury that has lasting effects, applies only to milder forms of neural tube defects and does not apply to anencephaly.
Most infants with anencephaly are easily diagnosed. Whenever cerebral remnants are visible through a defect in calvaria and skin, the diagnosis can be made; however, confusion may still result because infants with extreme microcephaly (ie, atelencephaly or aprosencephaly), microcephaly with an encephalocele covered by thin, nearly transparent fetal skin, or craniocerebral damage resulting from amniotic bands can be mistakenly diagnosed as anencephalic (166).
Iniencephaly. Iniencephaly is a rare, complex, and lethal form of neural tube defect that involves the occiput and inion, producing a characteristic extreme retroflexion of the neck.
Iniencephaly is more common among female infants. The cranium is closed and covered in skin (unlike anencephaly). The neck is severely retroflexed with an upward-looking face. The neck is short or may appear to be missing (due to fusion of cervical and thoracic vertebrae). The spine is usually closed but may be open in the cervical and thoracic regions. Variable features may include hydrocephalus, an occipital encephalocele, and a myelomeningocele or a meningocele.
Iniencephaly may be difficult to diagnose antenatally by ultrasound as it can be confused with other defects involving the brain and spine (eg, anencephaly or craniorachischisis, especially with neck retroflexion). Even postnatally, iniencephaly can be confused with craniorachischisis with spinal retroflexion.
Iniencephaly is a lethal condition. Iniencephaly is often (84%) associated with other findings, including certain chromosomal abnormalities (ie, trisomy 13, trisomy 18, and monosomy X) and other unrelated birth defects (eg, micrognathia, cleft lip and palate, cardiovascular disorders, diaphragmatic hernias, and gastrointestinal malformations).
The aprosencephaly/atelencephaly spectrum. In vertebrates, the forebrain (prosencephalon), the midbrain (mesencephalon), and hindbrain (rhombencephalon) are the three primary brain vesicles during the early development of the nervous system, listed from rostral to caudal. In humans, these divisions are evident by 5 weeks in utero. At the five-vesicle stage, at 8 weeks in utero, the forebrain separates into the diencephalon (thalamus, hypothalamus, subthalamus, and epithalamus) and the telencephalon, which develops into the cerebrum (cerebral cortex, underlying white matter, and basal ganglia).
The aprosencephaly/atelencephaly spectrum includes rare CNS malformations characterized by varying degrees of absence or dysplasia of the derivatives of the prosencephalon (ie, telencephalon and diencephalon), with an intact cranial vault: atelencephaly, in which only the telencephalon is affected, is the less severe form, whereas both the telencephalon and diencephalon are involved in aprosencephaly.
As summarized by Pasquier and colleagues, the aprosencephaly/atelencephaly spectrum
|
...refers to a rare forebrain malformation defined by a lack of prosencephalic derivates. In aprosencephaly the prosencephalon is absent, while in atelencephaly it exists as a rudimentary medial vesicle resembling the diencephalon but without the lateral telencephalic vesicles. Holoprosencephaly ... results from incomplete cleavage of telencephalic vesicles. ... [C]raniofacial malformations [in all of these disorders] involve the fronto-nasal eminence (ranging from cyclopia/synophthalmia [ie, fusion of both eyes in a single, central orbit located in the middle of the face], cebocephaly [ie, a developmental anomaly characterized by a monkey-like head with a small, flattened nose with a single nostril or absent nose and closely set eyes], and midline cleft to mild [orbital] hypotelorism or normal face) | |
|
(262) modified by Douglas Lanska |
Hence, aprosencephaly is the absence of the forebrain and its derivative structures, atelencephaly is the absence of the telencephalon (leaving only a rudimentary medial diencephalic structure), and holoprosencephaly is a failure of cleavage of the telencephalon so that the embryonic forebrain fails to divide into two lobes. During embryonic development, aprosencephaly is thought to occur after the optic vesicles form but before the cerebral vesicles appear (133). Atelencephaly and aprosencephaly differ from anencephaly in that the cranial vault is covered with skin, cerebral damage probably occurring by means of an encephaloclastic process after neural tube closure (319). Aprosencephaly/atelencephaly spectrum disorders are exceedingly rare, with approximately only 20 cases reported.
The amniotic band syndrome. Amniotic band syndrome occurs when the lining of the amniotic sac is damaged during pregnancy. This creates fibrous strands of tissue in which the fetus gets tangled. These strands, called amniotic bands, may wrap around and attach to different parts of the developing body, including the head, causing damage from constriction, distortion, and avulsion or amputation. The signs and symptoms vary greatly depending on the areas of the body involved and may include cleft lip or palate, acalvaria, encephalocele, exencephaly, anencephaly, and even acephaly (182; 155; 41; 351; 159; 126; 387; 58; 62; 284; 108; 213; 178; 345; 80; 81; 304; 386; 383).
Of CNS anomalies diagnosed prenatally, nearly half are neural tube defects (203). Prenatal diagnosis is primarily accomplished by imaging and the finding of an elevated alpha-fetoprotein through screening. When used alone, imaging appears to be more successful in diagnosing neural tube defects than alpha-fetoprotein screening (243). Substantive differences in screening policies still exist worldwide (28). Although these two tests have been generally successful, a significant number of cases are still missed (373).
Ultrasonography. First-trimester ultrasonographic diagnosis of anencephaly is possible at routine nuchal translucency examination, as early as 8 to 10 weeks, permitting further workup or termination of pregnancy when desired (165; 199; 173; 316). Anencephaly can be reliably diagnosed at the routine 10- to 14-week ultrasound scan, provided a specific search is made for the sonographic features for this condition (165). First-trimester ultrasonography by an expert sonographer with appropriate training, together with the application of a standardized protocol, is essential for a high detection rate of neural tube defects during a scan performed at 11 and 14 weeks (207).
(Source: Shrestha HK, Koirala S, Shrestha I. Misoprostol induced expulsion of fetus following diagnosis of anencephaly on ultrasound: a case report. JNMA J Nepal Med Assoc 2021;59[236]:396-8. Creative Commons Attribution 4.0 In...
In the first trimester, the pathognomonic feature on ultrasound is acrania, with variably evident disruption or distortion of the developing brain, whereas, in the second trimester, the absence of most of the brain is evident in addition to acrania (165). Because ossification of the fetal cranium is not completed until the eleventh week of pregnancy, fetal acrania can only be diagnosed after the eleventh week of pregnancy (165; 116; 07; 336). Between the eleventh and fourteenth weeks of pregnancy, most of the ossification points are found in lateral parts of frontal and parietal bones, whereas calvarium ossification is not yet visible in the mid-sagittal cross-section (336). Therefore, an incorrect diagnosis may occur if the U.S. imagining covers only a sagittal cross-section of the fetal cranium.
A high percentage (89%) of fetuses with acrania have echogenic amniotic fluid; hence, it is used as a supportive marker of fetal acrania in the first trimester (34). The presence of abundant but abnormal brain tissue observed on ultrasonography helps distinguish isolated acrania from more common anencephaly (202).
The main diagnostic 2D-ultrasound features of anencephaly may be characterized by (1) findings of acrania, (2) increased amniotic fluid echogenicity, and (3) the appearance in coronal images of a "Mickey Mouse face" (or a “Mickey-Mouse bi-lobular face") or a “frog face” or “frog eyes” sign (242; 59; 102; 207).
Twin pregnancies. In twin pregnancies, discordant amniotic fluid echotexture may be the first sign of the "acrania-exencephaly-anencephaly sequence" (207).
The "acrania-exencephaly-anencephaly sequence" for secondary anencephaly. Acrania itself can potentially transform into a secondary form of anencephaly (ie, as distinct from primary anencephaly due to failure of closure of the neural tube) through the so-called "acrania-exencephaly-anencephaly sequence" (369; 211; Cacifi and Sepulveda 2003; 127; 15; 207). Cerebral tissue that is not protected by the meninges, cranial bones, and skin may be gradually destroyed due to exposure to the harmful effect of amniotic fluid (eg, increased urea concentration in the amniotic fluid) and mechanical injuries (eg, friction with the uterine wall, placenta, and fetal limbs). Thus, acrania may lead to exencephaly—a lethal disorder of the fetal brain characterized by the absence of the calvarium and various degrees of remaining fetal brain tissue—and then ultimately to secondary anencephaly (Cacifi and Sepulveda 2003; 15).
In cases of the acrania-exencephaly-anencephaly sequence, first-trimester ultrasound shows a normal amount of brain tissue visible in the frontal plane of the fetus, resembling the rounded ears of Mickey Mouse—the “Mickey Mouse sign” or “Mickey Mouse face"—a sign typical for exencephaly. In the second trimester, a significant amount of brain tissue disappears, which manifests itself on ultrasonography as the “frog face” or “frog eyes” sign due to a lack of recognizable brain tissue above the fetal orbits—a sign typical for anencephaly (242; 59; 102).
Destroyed brain tissue suspended in small fragments in the amniotic fluid significantly increases the echogenicity of the amniotic fluid in such cases (211; Cacifi and Sepulveda 2003; 127).
In a series of five cases in the acrania-exencephaly-anencephaly sequence, cerebral structures were found on ultrasonography to be enclosed by a thin, inertial rippled membrane with a smooth outer contour (336). A thin anechoic space corresponding to CSF was observed between this membrane and underlying brain structures—the “beret sign." In three cases diagnosed with exencephaly, (1) the calvarium was absent; (2) brain structures had an irregular appearance with an unequal outer contour and remained motionless; and (3) amniotic fluid was anechoic. In two cases diagnosed with anencephaly, (1) the calvarium, meninges, and brain structures were missing, and (2) the “frog eyes” sign and slightly echogenic amniotic fluid were visible.
Scan performed using the transabdominal probe in 2D rendering in a 35-year-old gravida 3, para 1 woman at 12 weeks gestational age. Sagittal cross-section. Ultrasound diagnosis was acrania, confirmed at autopsy. Amniotic fluid ...
Scan performed using the transabdominal probe in 2D rendering in a 37-year-old gravida 4, para 2 woman with prior Cesarean section at 13 weeks gestational age. Ultrasound diagnosis was acrania, confirmed at autopsy. Sagittal cr...
Scan performed using the transabdominal probe in 2D rendering in a 25-year-old gravida 1, para 0 woman with prior Cesarean section at 13 weeks gestational age with cytomegalovirus infection. Ultrasound diagnosis was acrania, co...
Scan performed using the transabdominal probe in 3D rendering in a 25-year-old gravida 1, para 0 woman with prior Cesarean section at 13 weeks gestational age with cytomegalovirus infection. Ultrasound diagnosis was acrania, co...
Scan performed using the transabdominal probe in 2D rendering in a 30-year-old gravida 3, para 0 woman with diabetes at 13 weeks gestational age. Ultrasound diagnosis was acrania, but autopsy at 32 weeks showed anencephaly. Sag...
Scan performed using the transabdominal probe in 3D rendering in a 30-year-old gravida 3, para 0 woman with diabetes at 13 weeks gestational age. Ultrasound diagnosis was acrania, but autopsy at 32 weeks showed anencephaly. Sag...
Scan performed using the transvaginal probe in 2D rendering in a 21-year-old gravida 1, para 0 woman at 13 weeks gestational age. Ultrasound diagnosis was acrania, confirmed at autopsy. Sagittal cross-section. Amniotic fluid an...
Scan performed using the transvaginal probe in 3D rendering in a 21-year-old gravida 1, para 0 woman at 13 weeks gestational age. Ultrasound diagnosis was acrania, confirmed at autopsy. Sagittal cross-section. Amniotic fluid an...
Scan performed using the transvaginal probe in 2D rendering in a 21-year-old gravida 1, para 0 woman at 13 weeks gestational age. Ultrasound diagnosis was acrania, confirmed at autopsy. Coronal cross-section. Amniotic fluid ane...
Scan performed using the transvaginal probe in 2D rendering in a 22-year-old gravida 1, para 0 woman at 13 weeks gestational age. Ultrasound diagnosis was exencephaly, confirmed at autopsy. Sagittal cross-section. Cranial sonog...
Scan performed using the transabdominal probe in 2D rendering in a 29-year-old gravida 3, para 1 woman at 12 weeks gestational age. Ultrasound diagnosis was exencephaly, confirmed at autopsy. Sagittal cross-section. Cranial son...
Scan performed using the transvaginal probe in 2D rendering in a 24-year-old gravida 2, para 1 woman at 16 weeks gestational age. Ultrasound diagnosis was exencephaly, confirmed at autopsy. Sagittal cross-section. Cranial sonog...
Scan performed using the transabdominal probe in 2D rendering in a 25-year-old gravida 1, para 0 woman at 16 weeks gestational age with cytomegalovirus infection. Ultrasound diagnosis was anencephaly, confirmed at autopsy. Coro...
Scan performed using the transabdominal probe in 3D rendering in a 25-year-old gravida 1, para 0 woman at 16 weeks gestational age with cytomegalovirus infection. Ultrasound diagnosis was anencephaly, confirmed at autopsy. Sagi...
Scan performed using the transabdominal probe in 2D rendering in a 20-year-old gravida 1, para 0 woman at 25 weeks gestational age. Ultrasound diagnosis was anencephaly, confirmed at autopsy. Coronal cross-section. Cranial sono...
Three-dimensional ultrasonography. Three-dimensional ultrasonography, particularly with the HDlive rendering method, may confirm or further elucidate diagnoses made by 2-dimensional techniques (140; 207; 334; 336). The HDlive rendering method can generate amazingly realistic images of the human fetus from sonographic data using an advanced virtual illumination model and skin rendering techniques with the latest generation of beam-forming technology, speckle reduction algorithms, and compound resolution imaging technologies.
As a model case to show the potential of prenatal ultrasonography, a 28-year-old primigravida woman was referred for further evaluation of a suspected fetal cranial abnormality at 17 weeks of gestation (334). The medical and family history was unremarkable, and there was no history of drugs or substance abuse. 2D ultrasound images revealed the absence of calvarium with deformed brain tissue directly exposed to the amniotic cavity, suggestive of exencephaly. The fetal spine showed the absence of vertebral posterior elements with splaying of the lamina at the thoracic level. The lower limbs appeared to be fused in fixed extension with two femora and two tibiae, although it was unclear whether there were one or two feet. 3D rendering images using 4D ultrasound with HDlive mode showed the absence of a calvarium with deformed and degenerated brain tissues, consistent with exencephaly, as well as an absent right upper limb. The spine showed rachischisis with extensive clefts in the posterior part of the fetal vertebrae from the upper cervical region to the sacrum. Collectively, these findings were consistent with those of craniospinal rachischisis totalis (anencephaly with rachischisis totalis). In addition, the lower extremities were completely fused, and the feet were fused with the heels, which were immobile. The pregnancy was terminated at 18 weeks of gestation. External examination of the fetus revealed anencephaly with craniorachischisis totalis, with fused lower limbs (sirenomelia), absence of the right upper limb and external genitalia, and imperforate anus, closely matching the findings on the 3D sonographic rendering images. Radiographic imaging at autopsy demonstrated complete rachischisis: the fused lower limb contained two femora and two tibiae.
Two-dimensional ultrasound images at 17 weeks of gestation. Absence of vertebral posterior elements with splaying of the lamina at the thoracic level (white arrowhead). (Source: Sugiura T, Sato Y, Nakanami N, Tsukimori K. Prena...
The lower limbs appeared to be fused together in fixed extension with two femora and two tibiae (white dashed arrows). (Source: Sugiura T, Sato Y, Nakanami N, Tsukimori K. Prenatal sonographic image of sirenomelia with anenceph...
Three-dimensional HDlive rendered images at 17 weeks of gestation showing an extensive spinal cleft in the posterior part of the fetal vertebrae from the upper cervical region to the sacrum (white arrowheads). (Source:...
Fetal MRI. Fetal MRI has also been used with success (310; 35) and may be especially helpful in cases where prenatal ultrasound diagnosis is inconclusive (311).
Alpha-fetoprotein. In general, amniotic fluid alpha-fetoprotein is reliable as a screening test for anencephaly at and after 14 weeks. Maternal serum alpha-fetoprotein is most reliable at 16 weeks to 18 weeks. Elevations in maternal serum alpha-fetoprotein occur at the fetal-maternal interface and, therefore, amniotic fluid alpha-fetoprotein is not uniformly elevated in anencephaly. The finding must be evaluated carefully and supplemented with imaging studies, as other factors also result in elevated maternal serum alpha-fetoprotein (eg, gestational age, number of fetuses, maternal weight, race, presence of diabetes, or placental abnormality) (340). The combination of elevated maternal serum alpha-fetoprotein and low estriol appears to be especially predictive of anencephaly (382).
The marginal benefit of alpha-fetoprotein in addition to imaging may be quite low. Consequently, routine measurement of amniotic fluid alpha-fetoprotein during amniocentesis may not be warranted in centers with expertise in targeted ultrasonographic imaging, although medicolegal concerns may preclude substantive change in practice (320).
Postnatal diagnosis of anencephaly. The postnatal diagnosis of anencephaly is made by physical examination. No tests are necessary.
In rare cases, radiography may help further elucidate cranial anomalies. For example, a case of anencephaly with sirenomelia and renal agenesis was diagnosed by prenatal ultrasound, confirmed by postnatal radiography, and diagnosed as axial mesodermal dysplasia syndrome (339). Autopsy may also be helpful in identifying anomalies not discernible by imaging (55; 175).
Management is limited, as most liveborn anencephalics die soon after birth. The median survival time was 55 minutes in one study, although survival of 10 weeks has been reported (269). Comfort care for the affected infant and parental (and even staff) counseling is necessary (249).
The ethics of transplanting organs from anencephalic infants has long been debated without resolution (219; 214; 261; 222). Families should be counseled regarding this possibility (162) or the opportunity to donate tissue for research. Formal procedures have been established for this purpose (296; 10). Treatment of patients with other neural tube defects is, of course, highly diverse (123). Regardless of the nature of the defect, careful perinatal counseling is indicated (251).
Pregnancy in mothers carrying anencephalic fetuses is complicated by polyhydramnios, shoulder dystocia, and an increased fetal death rate (249). In cases of discordant monoamniotic twin pregnancy, the anencephalic twin may pose a risk to the survival of the healthy co-twin. In such cases, selective termination has been performed (305). Parents will be expected to sustain considerable grief over the loss of one twin and benefit from compassionate perinatal palliative care (218).
Anencephaly is a lethal disorder, and affected individuals are highly unlikely to undergo procedures that would require anesthesia.
All contributors' financial relationships have been reviewed and mitigated to ensure that this and every other article is free from commercial bias.

Douglas J Lanska MD MS MSPH
Dr. Lanska of the University of Wisconsin School of Medicine and Public Health and the Medical College of Wisconsin has no relevant financial relationships to disclose.
See ProfileNearly 3,000 illustrations, including video clips of neurologic disorders.
Every article is reviewed by our esteemed Editorial Board for accuracy and currency.
Full spectrum of neurology in 1,200 comprehensive articles.
Listen to MedLink on the go with Audio versions of each article.
MedLink®, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Developmental Malformations
Sep. 22, 2024
Epilepsy & Seizures
Sep. 06, 2024
Developmental Malformations
Aug. 11, 2024
Developmental Malformations
Aug. 06, 2024
Developmental Malformations
Aug. 05, 2024
General Child Neurology
Aug. 05, 2024
Developmental Malformations
Aug. 03, 2024
Developmental Malformations
Jul. 06, 2024