Sleep Disorders
Sudden infant death syndrome
Jul. 05, 2026
MedLink, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Worddefinition
At vero eos et accusamus et iusto odio dignissimos ducimus qui blanditiis praesentium voluptatum deleniti atque corrupti quos dolores et quas.
Sleep and its disorders are complex, and their mechanisms remain only partially understood. However, neuroimaging studies investigating specific aspects of several sleep disorders have shown that functional as well as structural brain alterations can be identified. Structural and functional alterations can inform us of the neural mechanisms underlying the nighttime and daytime manifestations of sleep disorders, including their impact on cognitive function. Therefore, neuroimaging may assist us in uncovering the physiological underpinnings of these conditions and the targets for the development or monitoring of sleep-related interventions. There is a wide range of sleep disorders, and the following sleep disorders are discussed in this article: insomnia, narcolepsy and idiopathic hypersomnia, REM sleep behavior disorder, and obstructive sleep apnea.
|
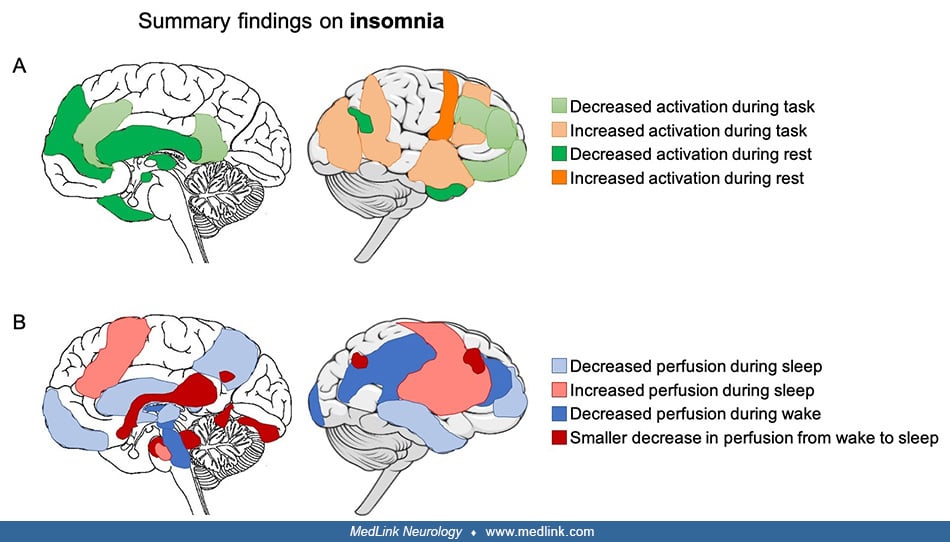
• In insomnia, a variety of cognitive and emotional tasks have been used to evaluate functional brain alterations. They generally point to a hyperactivation in response to sleep-related and negative emotional stimuli and a hypoactivation in response to other types of stimuli, such as tasks involving executive control. In contrast, structural brain alterations are inconsistent, perhaps reflecting the complexity and heterogeneity of insomnia and suggesting that sleep-related symptoms in insomnia are reflected in functional rather than structural brain alterations. | |
|
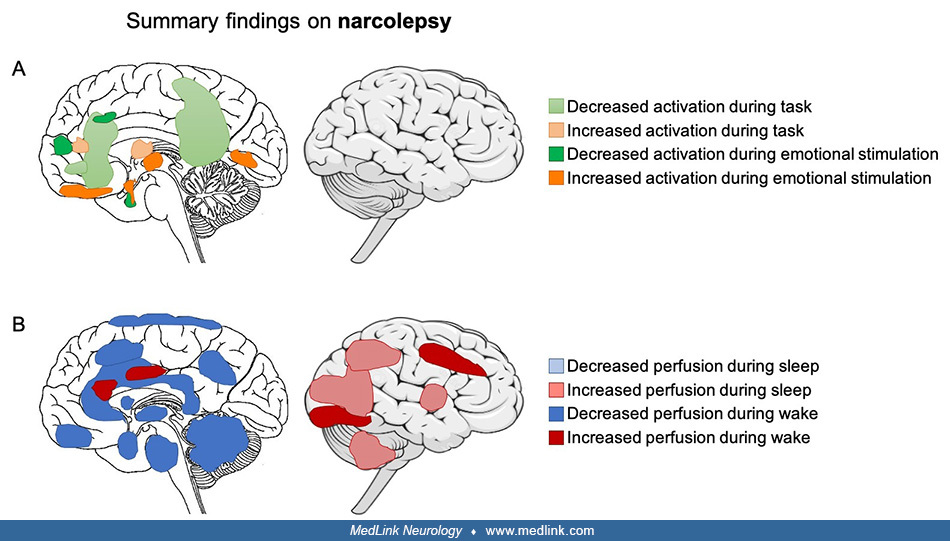
• In narcolepsy with cataplexy, structural alterations, such as smaller gray matter volume and altered white matter integrity, are widespread and encompass the hypothalamus and its projections, which is in line with the evidence for loss of hypocretin neurons in the hypothalamus. On the other hand, functional alterations are observed in the hypothalamus, which shows altered activation and regional blood flow during both resting wakefulness and while performing cognitive tasks, such as emotion regulation during positive emotions or cataplexy attacks. | |
|
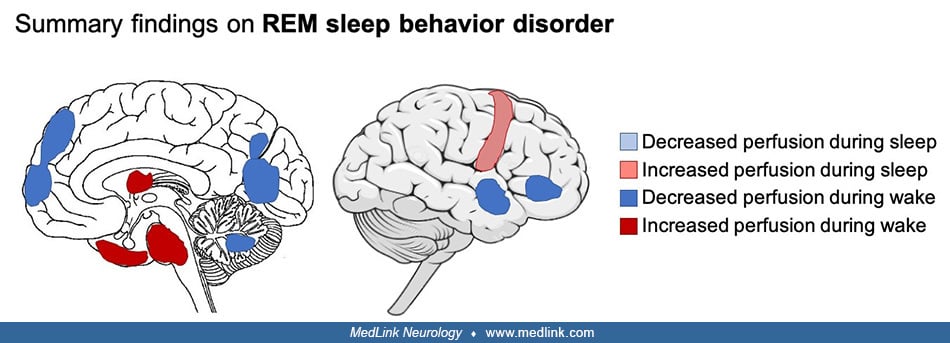
• In REM sleep behavior disorder, functional alterations are widespread across the cortex and involve the limbic network, pons, and thalamus. Specifically, the basal ganglia and sensorimotor network show altered functional connectivity, which might be a predictor of disease progression from REM sleep behavior disorder to Parkinson disease. | |
|
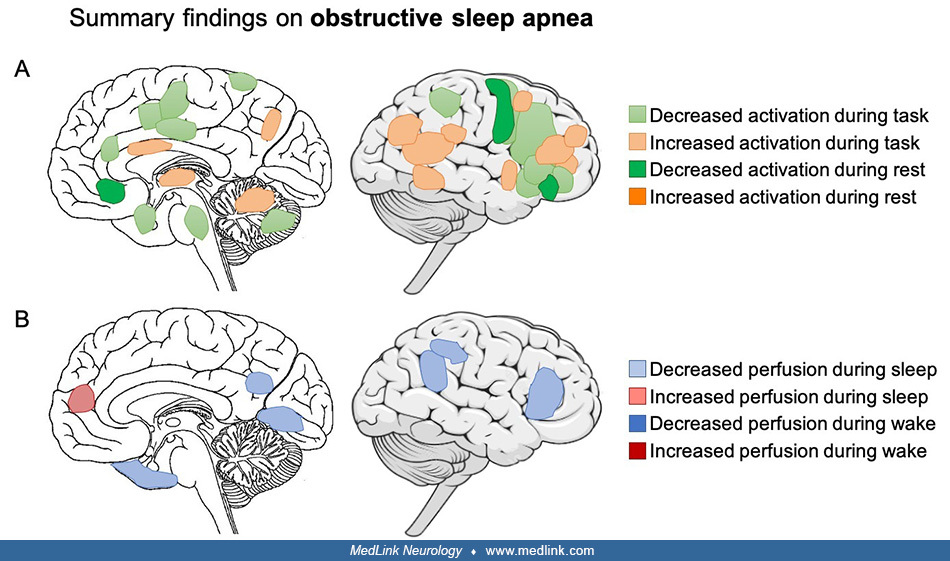
• In obstructive sleep apnea, cognitive deficits might stem from lower resting state activity and functional connectivity in the frontoparietal, sensorimotor, and salience networks, which are involved in numerous cognitive processes. However, the pattern of structural alterations is less consistent. Both structural and functional alterations may be partially reversible with continuous positive airway pressure treatment. |
Important advances in the understanding of sleep and its disorders have been made using neuroimaging techniques, such as magnetic resonance imaging (MRI), positron emission tomography (PET), or single-photon emission computed tomography (SPECT), as they bring complementary information to electroencephalography (EEG). The understanding of sleep physiology and sleep disorders may, therefore, be improved by investigating structural and functional brain alterations during resting wakefulness, sleep, or while performing a task using these neuroimaging techniques.
MRI is used in clinical practice to investigate brain anatomy, but structural MRI can also provide measures of cortical thickness, regional gray or white matter volume, white matter integrity, and structural connectivity by looking at how water molecules diffuse in the brain. Functional MRI (fMRI) measures changes in cerebral blood flow through a blood-oxygen-level-dependent contrast. Regional brain activity is indirectly measured by measuring the increase of oxygen consumption and, therefore, blood flow resulting from neuronal activity. fMRI can also be used to assess brain activity patterns and functional connectivity. Brain activity can be measured when subjects perform a cognitive task, during sleep, or during resting wakefulness (ie, in the absence of any particular task or sleep). Functional connectivity looks at coactivated brain regions, ie, how different brain regions or networks may communicate with each other. Finally, with magnetic resonance spectroscopy (MRS), one can measure metabolites or neurotransmitter concentration in a specific brain region. Metabolite levels that can be measured include gamma-aminobutyric acid (GABA) and glutamate to assess excitatory and inhibitory neurotransmission. Other molecules can be measured and are used as markers of neuronal or glial integrity, such as N-acetylaspartate, choline, and creatine. In most cases, metabolite levels are measured relative to another metabolite as metabolites are difficult to quantify individually.
There are two main types of nuclear imaging techniques. The first type is positron emission tomography (PET) imaging and involves injecting a radioactive tracer into the bloodstream, which is detectable within the brain. Depending on the tracer, it is possible to measure the density of neurotransmitter receptors or the regional brain metabolic rate. For example, [18F]-fluorodeoxyglucose positron emission tomography (FDG-PET) provides a measurement of regional glucose metabolism within the brain over a short timespan (eg, 20–30 min). The second type is single-photon emission computed tomography (SPECT), a technique that also utilizes the injection of a tracer into the bloodstream and can be used to quantify regional cerebral perfusion (blood flow, rCBF) or neurotransmission (depending on the tracer used) in the brain.
|
• In insomnia, brain anatomy alterations in the frontoparietal network have been reported but are not consistent across studies, perhaps reflecting the complexity and heterogeneity of insomnia disorder and suggesting that sleep-related symptoms in insomnia are reflected in functional rather than structural brain alterations. | |
|
• Insomnia is characterized by a hyperactivation in response to sleep-related and negative emotional stimuli and a hypoactivation in response to other types of stimuli, such as tasks involving executive control. | |
|
• Brain activity and connectivity during resting wakefulness is altered in insomnia, particularly within the default mode and salience networks and between the limbic network and other brain regions. These alterations may contribute to the state of hyperarousal and emotional dysregulation that are present in insomnia. | |
|
• The overall activity decrease in the default mode network in the transition from wakefulness to sleep is less pronounced in insomnia and might reflect heightened arousal. |
Definition of insomnia. Insomnia is characterized by difficulties falling asleep and maintaining sleep as well as waking up earlier than desired. It is associated with daytime function impairments, such as fatigue and reported cognitive impairments. The distinguishing features of insomnia have been associated with functional alterations that are, for the most part, in line with the hyperarousal theory of insomnia, which postulates that heightened physiological, cognitive, or emotional arousal observed in insomnia is the result of excessive central nervous system arousal that prevents and disturbs sleep.
Functional brain alterations during cognitive tasks in insomnia. Various cognitive and emotional tasks have been used to evaluate functional brain alterations in insomnia and generally point to a hyperactivation in response to sleep-related and negative emotional stimuli and a hypoactivation in response to other types of stimuli, such as tasks involving executive control (45; 47; 55; 73; 85).
The relationship between emotion and hyperarousal has been investigated in several fMRI studies. In a study, greater activation in response to emotional stimuli was present in the amygdala in individuals with insomnia when trying to reinterpret the meaning of negative pictures (23). However, this hyperarousal might be specific to sleep and insomnia as only emotionally arousing stimuli that were related to sleep elicited greater amygdala activation in an emotional reactivity task in individuals with insomnia (05). In addition, the habituation to emotional images that was observed in good sleepers over successive trials was absent in individuals with insomnia (05).
The impairment of daytime functioning is a key feature of insomnia and mainly affects subjective cognitive functioning. Despite inconsistent evidence for objective cognitive impairments, brain cognitive processing as shown by fMRI seems to be altered. Individuals with insomnia had reduced activation of the left medial and inferior frontal gyri during a verbal fluency task involving the executive control network, without showing poorer performance (03). During a working memory task, lower activation in the frontal gyrus and motor and cingulate cortices was observed in individuals with insomnia; they also failed to deactivate the default mode network (21).
Functional brain alterations during resting wakefulness in insomnia. The growing literature on resting wakefulness in insomnia shows an altered brain activity and connectivity, particularly within the default mode and the salience networks and between the limbic network and other brain regions, which are thought to contribute to hyperarousal, emotional regulation, and cognitive alterations that are present in insomnia. Resting state fMRI functional connectivity was found to be lower in the salience and executive control networks and higher in the sensorimotor network (45). In the salience network, the activation of which is associated with negative internal and external stimuli, functional connectivity was associated with negative effect in individuals with insomnia (15; 73). In the limbic network, functional connectivity was decreased between the thalamus and left amygdala, parahippocampal gyrus, putamen, pallidum, and hippocampus (85) as well as between the amygdala and the insula, striatum, and thalamus in individuals with insomnia (37). The latter study also found increased connectivity between the amygdala and the premotor and sensorimotor cortices, which was associated with decreased sleep quality. Altered functional connectivity within the limbic network may be reflective of altered emotional processing in insomnia, and the increased functional connectivity between the amygdala and sensorimotor cortices may support a model of excessive hyperarousal. Additionally, altered functional connectivity between the locus coeruleus, which is involved in the transition between sleep and wake, and other brain networks was associated with poorer sleep quality and greater insomnia symptoms (26; 52).
In the default mode network, a large study from the UK Biobank reported an association between frequent insomnia symptoms (n = 9210) and increased connectivity within the default mode network (33). In addition, an increased negative connectivity between the default mode network and the frontoparietal network, and decreased connectivity between the salience network and a node of the default mode network, were reported. However, inconsistent alterations of functional connectivity and regional brain activity in the cingulate cortex, left insula, left cuneus, and left fusiform gyrus were also observed in smaller studies. Indeed, some studies showed increased connectivity and activation in the default mode network, whereas other showed decreased activity (45). Altered functional connectivity in the default mode network was often associated with subjective sleep disturbances. Additionally, decreased functional connectivity between the medial prefrontal cortex and middle temporal lobe and between the left temporal lobe and left inferior parietal cortex was found in individuals with insomnia (60). Because these regions play an important role in cognitive functioning and sleep physiology, these findings suggest that decreased connectivity may underlie the alteration in cognitive functioning, whereas increased activity at rest might reflect a hyperarousal state in individuals with insomnia. The reduction in brain activation during resting wakefulness has been confirmed using FDG-PET, showing an overall reduction in glucose metabolism in individuals with insomnia. More specifically, an altered engagement of the limbic, default mode, and executive control networks might be reflected by lower middle frontal, fusiform, lingual gyri, precuneus/posterior cingulate, and frontoparietal cortex glucose metabolism in insomnia (46).
Functional brain alterations during sleep and sleep-wake transitions in insomnia. Significant brain activity changes occur during sleep: the overall level of activity decreases in the default mode network, and there is a disconnection (functional connectivity decrease) between its posterior and anterior parts. More specifically, the posterior cingulate cortex and inferior parietal lobe disconnect from the medial prefrontal cortex and anterior cingulate cortex during sleep (35). This disconnection becomes more pronounced as sleep deepens and reaches its lowest level during slow-wave sleep. In individuals with insomnia, this overall activity decrease that happens during sleep seems less pronounced as the decline in regional brain glucose metabolism from wakefulness to sleep is smaller in the medial prefrontal cortex, anterior cingulate, insula, thalamus, hippocampus, amygdala, hypothalamus, and brainstem (61). This was confirmed by another FDG-PET study in a larger sample of individuals showing a smaller decline in brain glucose metabolism during NREM sleep in the default mode network (46). Alterations in other networks are present during sleep in individuals with insomnia as fMRI functional connectivity between the thalamus and left amygdala, parahippocampal gyrus, putamen, pallidum, and hippocampus was decreased across all threec NREM sleep stages.
Structural brain alterations in insomnia. Brain anatomy alterations are inconsistent in individuals with insomnia, potentially because insomnia is a complex and heterogeneous disorder. The discrepancy of results as well as the lack of large sample data make it difficult to draw a clear picture of structural alterations in insomnia and suggest that sleep-related symptoms in insomnia are reflected in functional brain alterations rather than structural brain alterations. A large sample study of 1053 patients with major depressive disorder from the Enhancing NeuroImaging Genetics through Meta-Analysis (ENIGMA) major depressive disorder consortium showed that insomnia severity was specifically related to smaller cortical surface area in the right insula, left inferior frontal gyrus pars triangularis, left frontal pole, right superior parietal cortex, right medial orbitofrontal cortex, and right supramarginal gyrus and was independent of major depressive disorder (51). In addition, cortical thinning was found in the anterior cingulate cortex, precentral cortex, and lateral prefrontal cortex in individuals with insomnia disorder (76). However, a subsequent study from the ENIGMA-Sleep consortium did not report any insomnia-specific brain alterations or any significant associations between insomnia symptoms and cortical or subcortical volumes (82).
Previous smaller sample studies had inconsistent findings regarding gray matter volume. Hippocampal gray matter volume was found to be reduced in individuals with insomnia in one study, but this finding was not replicated in other studies. The prefrontal regions (orbitofrontal and dorsolateral prefrontal cortex) were found to be smaller in individuals with insomnia in two studies, but this was not replicated in a latter study. Increased anterior cingulate cortex gray matter volume was found to be associated with increased insomnia severity in individuals with insomnia, but this was not replicated in a larger study.
Fewer studies have examined white matter alterations. Overall, white matter integrity was affected in frontostriatal, frontothalamic, and cortico-cortical networks as well as within the limbic network (mostly shown by reductions in fractional anisotropy or less restricted water diffusion) in individuals with insomnia (59). Lower tract strength, as a marker of poorer white matter integrity, was observed within the limbic network and was negatively correlated with sleep quality in individuals with insomnia (14), showing the importance of emotion regulation in insomnia.
Brain metabolite alterations in insomnia. In terms of neurochemistry, the main metabolite that has been investigated in insomnia is GABA, which is involved in sleep-wake regulation as demonstrated by the use of benzodiazepines (positive allosteric modulators of the GABAA receptor) for the short-term treatment of insomnia. Individuals with insomnia may present an overall reduced inhibitory tone as lower GABA levels have been found in the anterior cingulate and in parieto-occipital cortices. However, higher GABA levels have been found in the occipital cortex (45). Only a handful of studies have investigated other metabolite levels, without consistent results. Phosphocreatine was decreased in individuals with insomnia, suggesting altered cortical energetic demands (30). Aspartate, glutamine, and creatine were decreased in the lateral occipital cortex in individuals with insomnia and short sleep duration (57). Glutamate, an excitatory neurotransmitter, was increased in the parieto-occipital cortex and was associated with insomnia symptoms in patients with posttraumatic stress disorder (45).
|
• In narcolepsy with cataplexy, structural alterations, such as smaller gray matter volume and altered white matter integrity, are widespread and encompass the hypothalamus and its projections, which is in line with the evidence for loss of hypocretin neurons in the hypothalamus. | |
|
• Functional alterations in narcolepsy with cataplexy are observed in the hypothalamus, which shows altered activation and regional blood flow during both resting wakefulness and while performing cognitive tasks, such as emotion regulation during positive emotions or cataplexy attacks. | |
|
• Idiopathic hypersomnia does not show similar structural alterations as compared to narcolepsy with cataplexy but seems to present a disruption of the default mode network, both at the structural level and at the level of functional connections within regions of the default mode network. |
Definition of narcolepsy and idiopathic hypersomnia. Narcolepsy is characterized by excessive daytime sleepiness and abnormalities in REM sleep. Narcolepsy is categorized into two types: narcolepsy with cataplexy (type 1) and narcolepsy without cataplexy (type 2). Narcolepsy with cataplexy is associated with a deficiency in the hypothalamic neuropeptide orexin-A (hypocretin-1), which is involved in the abnormal sleep-wake patterns and cataplexy that are characteristic of narcolepsy. Idiopathic hypersomnia is primarily characterized by excessive daytime sleepiness, together with an often-prolonged total sleep time and difficulties waking up (sleep drunkenness), for which the pathophysiological mechanisms remain unclear. Individuals suffering from idiopathic hypersomnia do not present cataplexy, rapid REM sleep onset (at the multiple sleep latency test), or any consistent hypocretin-1 deficiency.
Functional brain alterations during cognitive tasks in narcolepsy. Based on clinical observations that cataplectic episodes in narcolepsy can be triggered by positive emotions, brain processing involved in emotion and reward have been studied (16). Two fMRI studies used humorous images to study emotional processing and showed alterations in the amygdala and hypocretin system. The first study found reduced hypothalamic activity and increased amygdala activity (74). The second study found enhanced activity in the hypothalamus and emotional network (67). Reward processing was altered in individuals with narcolepsy as they failed to show functional connectivity between the amygdala and medial prefrontal cortex (65). The investigation of other tasks, such as attention or working memory, suggest that narcolepsy is characterized by an imbalance of cognitive resources allocation in favor of monitoring and maintaining attention (27).
Functional brain alterations during resting wakefulness in narcolepsy. Investigation of resting wakefulness in narcolepsy suggests decreased activity in regions that are important targets of hypocretinergic projections (16). An example is decreased regional cerebral blood flow in the brainstem in individuals with narcolepsy (56). Another study found decreased perfusion in the hypothalamus, caudate nuclei, thalamus, cingulate gyrus, and frontoparietal cortex in individuals with narcolepsy with cataplexy (42). However, there have been inconsistent findings regarding glucose metabolism: a first study found hypometabolism in the bilateral hypothalami, thalamus, and frontal-parietal cortex (44) in individuals with narcolepsy, whereas a second study only found hypermetabolism in the cingulate and visual association cortices (19). Similarly, functional connectivity changes have been found in narcolepsy type 1. Increased functional connectivity was found in the left middle occipital gyrus, right inferior occipital gyrus, and lingual gyrus and between the amygdala and the inferior frontal gyrus, claustrum, insula, and putamen. In contrast, decreased functional connectivity was found in the posterior lobe of the cerebellum, left inferior temporal gyrus, and left dorsolateral superior frontal gyrus and between the lateral hypothalamus and the left superior parietal lobule, the hippocampus, and the parahippocampal gyrus, as well as between the amygdala and the post-central gyrus and several occipital regions (07). In addition, a new imaging technique investigating brain pulsation showed altered CSF dynamics in narcolepsy type 1, suggesting altered glymphatic process (41).
Functional brain alterations during sleep in narcolepsy. Findings during REM sleep are inconsistent: some studies report increased activity or an absence of decreased activity that is supposed to occur during sleep (16). A later study found decreased activity in the hypothalamus, caudate nuclei, thalamus, cingulate gyrus, and frontoparietal cortices in narcolepsy with cataplexy (42).
Functional brain alterations during cataplexy in narcolepsy. Because it is difficult to capture cataplectic episodes, only case studies have reported such results, and they show inconsistent findings. Hyperperfusion has been found in limbic areas (including the amygdala and orbitofrontal and anterior cingulate cortices), the thalamus, basal ganglia, brainstem, and parietal cortices (34), whereas hypoperfusion has been found in the prefrontal and occipital cortices (34) and precentral and primary somatosensory cortex (19). The latter observation was supported by an fMRI study that showed marked hypoactivation of the hypothalamus during a cataplectic attack (67).
Functional brain alterations in idiopathic hypersomnia. Only two neuroimaging studies have investigated functional alterations in idiopathic hypersomnia. Although these findings require replication, they suggest functional alterations in brain areas involved in the modulation of vigilance states rather than a loss of hypothalamic orexin neurons. One study found that glucose metabolism was increased in the insula, anterior and middle cingulate cortex, and caudate nucleus (20). Another study showed that regional cerebral blood flow was lower in the medial prefrontal cortex as well as in the posterior cingulate cortex, putamen, and cerebellum in individuals with idiopathic hypersomnia (10). Using fMRI, the same group showed lower functional connectivity within the anterior part of the default mode network (ie, medial prefrontal cortex), which was also associated with sleepiness (64).
Structural brain alterations in narcolepsy. Early structural studies of narcolepsy focused on the pontine tegmentum region in the brainstem as this structure controls the transition between vigilance states, but they found inconsistent results that may be due to heterogeneity in patient characteristics or neuroimaging techniques. Later, in line with evidence suggestive of a hypocretin deficiency, several studies investigated gray matter volume of the hypothalamus. Meta-analyses of those studies have shown inconsistent alterations of thalamic and hypothalamic structures in narcolepsy (28). However, consistent gray matter volume decreases in frontotemporal regions, which might be associated with cognitive complaints of memory and attention, have been observed in narcolepsy with cataplexy. Additionally, alterations in white matter integrity have been found in the hypothalamus-thalamus-orbitofrontal pathway and in the brainstem as well as in the inferior fronto-occipital fasciculus. However, no studies have specifically investigated structural brain alterations in narcolepsy without cataplexy.
Structural brain alterations in idiopathic hypersomnia. Fewer studies have investigated structural brain alterations in idiopathic hypersomnia, and those few reports show no global gray or white matter volume alterations. However, regional brain alterations have been found. Cortical thickness and gray matter volume are larger in the posterior default mode network and in the middle occipital gyrus in individuals with idiopathic hypersomnia. More specifically, the volume of the precuneus was positively associated with greater subjective daytime sleepiness (64).
Brain metabolite alterations in narcolepsy. The investigation of brain metabolite levels in narcolepsy suggests decreased N-acetylaspartate/creatine levels in the hypothalamic region, which is consistent with evidence that hypocretinergic neurons are selectively damaged in narcolepsy (77). However, the thalamus and parieto-occipital cortex, which are also innervated by those neurons, did not reveal any alterations in N-acetylaspartate/creatine levels (78). Finally, absolute GABA concentrations were found to be higher in the medial prefrontal cortex in individuals with narcolepsy, and this was positively associated with nocturnal sleep quality (49).
|
|
• Because of the link between REM sleep behavior disorder and Parkinson disease, structural alterations were primarily investigated in the pons and midbrain as well as in the basal ganglia, all of which involve dopamine transmission. However, structural alterations in REM sleep behavior disorder also encompass temporal, frontal, and parietal cortices. |
|
• Functional alterations are widespread across the cortex and involve the limbic network, sensorimotor network, pons, and thalamus. More specifically, alteration of functional connectivity within the basal ganglia may be an important marker of basal ganglia network dysfunction. |
Definition of REM sleep behavior disorder. REM sleep behavior disorder is a disorder characterized by the loss of muscle atonia during REM sleep, which leads to dream enactment that often results in dramatic movements, such as punching, flailing, or jumping out of the bed, as well as injuries and problems with the bed partner. There is increasing evidence that REM sleep behavior disorder is associated with the alpha-synucleinopathies, such as Parkinson disease, dementia with Lewy bodies, and multiple system atrophy; REM sleep behavior disorder is considered a risk factor for alpha-synucleinopathies (40).
Functional brain alterations in REM sleep behavior disorder. Decreased perfusion has been found in the frontal and temporoparietal cortices of individuals with REM sleep behavior disorder; decreased cerebral blood flow has been found in the parietal and occipital cortices as well as in the cerebellum and limbic network (16). Increased perfusion has been observed in the pons, putamen, and hippocampus. The hyperperfusion within the hippocampus predicted the development of dementia with Lewy bodies or Parkinson disease 3 years later, and this occurred in half of the patients (18). REM sleep behavior disorder, a risk factor for alpha-synucleinopathies (40), suggests a dysfunction in dopaminergic transmission, which might play a role in the process leading to neurodegeneration. In addition, decreased glymphatic system flow was associated with increased conversion risk to alpha-synucleinopathies (04). A few neuroimaging studies investigated the dopaminergic system and showed greater dopamine transporter density in individuals with REM sleep behavior disorder, whereas other studies showed no difference or lower dopamine transporter density (16). Longitudinal studies have shown that individuals who developed a neurodegenerative disorder had lower dopamine transporter at baseline (38).
In addition, functional connectivity is altered in the basal ganglia network (68) and in the motor cortex and sublaterodorsal tegmental nucleus (24) in individuals with REM sleep behavior disorder. Lower within-network functional connectivity has been identified between the striatal and prefrontal regions, within the executive control network, between the midbrain and pallidum, within the basal ganglia network, and between motor and somatosensory regions as well as within the sensorimotor network (81). Finally, global basal ganglia resting state connectivity in REM sleep behavior disorder was similar to individuals with Parkinson disease, suggesting that activity within the basal ganglia network may be a marker of basal ganglia dysfunction (68).
Structural brain alterations in REM sleep behavior disorder. REM sleep behavior disorder is characterized by smaller gray matter volume in the thalamus; pontomesencephalic tegmentum; and temporal, frontal, and parietal cortices (24). More severe reductions in volume have been found in Parkinson disease with associated REM sleep behavior disorder (50). Only a couple of studies found increased volume in the caudate nucleus (32). This lower volume in the pontomesencephalic tegmentum is of importance because it contains cholinergic, GABAergic, and glutamatergic neurons implicated in the promotion of REM sleep and muscle atonia (11). Idiopathic REM sleep behavior disorder has also been associated with higher perivascular space burden in the centrum semiovale, basal ganglia, substantia nigra, and brainstem compared to individuals with Parkinson disease, which correlates with REM sleep behavior disorder symptom severity (75). White matter alterations such as higher fractional anisotropy, a marker of more restricted water diffusion, have been found in the pons (80) and other regions known to be involved in REM sleep regulation, such as the pontine region, (72), cerebellar peduncles, and brainstem (32).
Brain metabolite alterations in REM sleep behavior disorder. REM sleep behavior disorder does not seem to involve detectable metabolite alterations in mesopontine regions. Only one case study revealed an increased choline/creatine ratio in the brainstem (58); others failed to replicate these findings (39).
|
• Cognitive deficits observed in obstructive sleep apnea might be explained by lower resting state activity and functional connectivity in the frontoparietal, sensorimotor, and salience networks, which are involved in numerous cognitive processes. | |
|
• In contrast, the pattern of structural alterations in obstructive sleep apnea is less consistent. Gray matter atrophy has been observed in the frontal and parietal cortex, temporal lobe, anterior cingulate, thalamus, hippocampus, and cerebellum. White matter alterations encompass the corpus callosum, sensorimotor cingulate cortex, corticospinal tract, insular cortex, basal ganglia, and limbic network. | |
|
• Functional and structural changes may be partially reversible with continuous positive airway pressure treatment. |
Definition of obstructive sleep apnea. Obstructive sleep apnea is characterized by recurrent episodes of partial or complete occlusion of the upper airway during sleep, which impedes breathing despite continued inspiratory efforts. The repetitive obstructive events that occur during sleep in obstructive sleep apnea result in variable changes in blood gas concentrations, which lead to hypoxemia (low blood oxygen) and hypercapnia (high blood carbon dioxide). Due to the body’s attempts to regain airflow, the apneic events are strongly correlated with arousals from sleep, leading to significant sleep fragmentation and a reduction in deeper sleep stages. It is well established that obstructive sleep apnea is associated with impairments in cognitive function, particularly in vigilance, attention, memory, and executive function domains. Importantly, obstructive sleep apnea can be controlled with treatments such as continuous positive airway pressure, which serves to maintain airway patency and prevent obstructions.
Functional brain alterations during cognitive tasks in obstructive sleep apnea. Because obstructive sleep apnea is associated with cognitive deficits, functional alterations are more easily detected while performing a cognitive task. A multimodal meta-analysis showed lower activation during various cognitive tasks in individuals with obstructive sleep apnea, specifically in the dorsolateral prefrontal and orbitofrontal cortices; higher activation was found in the insula, anterior cingulate cortex, amygdala, and hippocampus (36). This finding was based on small sample studies investigating different cognitive domains in which working memory, executive functioning, and attention performance were poorer in individuals with obstructive sleep apnea. Lower resting state functional connectivity in the sensorimotor network, salience (insula) network, and frontoparietal network and altered within-network coherence might explain the impaired cognitive and motor performance observed in individuals with obstructive sleep apnea (47). Continuous positive airway pressure treatment can reduce hypoxia and normalize blood oxygen saturation, thereby reducing sleep fragmentation and improving cognitive function. After continuous positive airway pressure treatment, restoration in abnormal baseline activation in the left inferior frontal gyrus, anterior cingulate cortex, and hippocampus (12) and an increase in task-related activation (66) were associated with a significant improvement in working memory performance.
Functional brain alterations during resting wakefulness in obstructive sleep apnea. Overall, studies show that there is a decrease in brain activity as indicated by decreased perfusion in the parietal cortex (22), bilateral parahippocampal gyri, and right lingual gyrus (43) in individuals with obstructive sleep apnea during resting wakefulness. Functional connectivity within dorsal and ventral attention networks as well as the default-mode network is decreased at rest, and this decrease is associated with worst cognitive performance (31). There is also decreased metabolism in the precuneus, middle and posterior cingulate gyrus, parieto-occipital cortex, and prefrontal cortex (84). These decreases might be due to compensatory mechanisms in response to blood flow alterations or blood gas changes in obstructive sleep apnea and are partly reversible with continuous positive airway pressure treatment (48; 54).
Structural brain alterations in obstructive sleep apnea. Structural alterations in obstructive sleep apnea might correspond to neuronal damage resulting from intermittent hypoxia. Reduced cortical thickness has been found in individuals with obstructive sleep apnea in the precentral and postcentral gyri, ie, sensorimotor areas, corresponding to the upper airway musculature areas as well as temporal cortex and insula (53). Cortical thinning in the hippocampus and entorhinal cortex has been found to be related to disease severity and can be slowed down by continuous positive airway pressure treatment (63). In contrast, other studies reported greater cortical thickness in frontal regions (08; 17; 25), which were associated with obstructive sleep apnea severity and could, therefore, be reflective of edema and reactive cellular processes (08).
Gray matter volume decreases seem to be more widespread and encompass the prefrontal, postcentral gyrus, temporal, and parietal cortices as well as the anterior cingulate cortex, posterior hippocampus (CA3/dentate), thalamus, and cerebellum. Most of these changes have been associated with cognitive deficits or disease severity, and smaller hippocampal volume has been associated with excessive daytime sleepiness (83; 36; 06). In contrast, increases in gray matter volume seem to be restricted to the CA1, subiculum, and uncus parts of the hippocampus (83).
White matter integrity is also affected in individuals with obstructive sleep apnea. Mean diffusivity is lower (ie, water diffusion is more restricted) in the corpus callosum, sensorimotor cingulate cortex, corticospinal tract, insular cortex, basal ganglia, and limbic network (13; 69) and might be reversible after treatment with continuous positive airway pressure (63). In some individuals with obstructive sleep apnea, white matter hyperintensities can be observed on MRI scans and might reflect vascular damage that is associated with aging or comorbid hypertension (69).
Brain metabolite alterations in obstructive sleep apnea. The cognitive deficits in obstructive sleep apnea may also be related to the metabolite alterations in specific cerebral regions. The level of N-acetylaspartate/creatine has been found to be decreased in periventricular and frontal white matter (16), possibly reflecting poorer neuronal integrity and glial dysfunction. In addition, choline/creatine levels were increased in the thalamus (01) and temporal cortex (71), which might indicate reactive gliosis or maladaptive changes in membrane metabolism in response to hypoxia. In contrast, other studies have found increased N-acetylaspartate/creatine and choline/creatine ratios in the hippocampus, which correlated with greater obstructive sleep apnea severity (09; 02). Finally, some metabolite ratio alterations were reversible with continuous positive airway pressure treatment (62), whereas others were not (79).
|
• Although structural alterations observed with neuroimaging can reflect evidence of a disturbance in specific neurotransmission systems (eg, REM sleep behavior disorder) or long-term consequences of a sleep disorder (eg, the consequences of hypoxia in obstructive sleep apnea), the structural and functional alterations are more subtle and less generalizable in other sleep disorders, such as insomnia. | |
|
• More large-scale studies and multisite datasets are needed to better identify the more subtle structural and functional alterations that are present in sleep disorders. | |
|
• Neuroimaging has generated valuable insights into the understanding of the neurologic bases and consequences of different sleep disorders, thereby helping to better delineate sleep disorders, refine diagnoses, and help in the development of treatment approaches. |
Neuroimaging can help track the progression of sleep disorders; for example, in REM sleep behavior disorder, the level of dopamine neurotransmission appears to be related to the progression of REM sleep behavior disorder. Nuclear imaging studies have also shown that lower dopamine receptor density is observed in subclinical REM sleep behavior disorder and later transitions to synucleinopathies, such as Parkinson disease and Lewy body dementia. However, it is unclear whether such dopamine abnormalities are a causal factor or consequence of REM sleep behavior disorder. In addition, resting state functional connectivity in the basal ganglia, compared to norms in a healthy population, could also be a marker of disease progression. It has been shown to be similar in REM sleep behavior disorder and in individuals with Parkinson disease; these findings need to be replicated in longitudinal studies (68).
Further, neuroimaging can help refine diagnoses more specifically for central hypersomnia disorders. Neuroimaging can be an additional tool to better delineate narcolepsy with cataplexy from narcolepsy without cataplexy or idiopathic hypersomnia. Neuroimaging can also shed light on the interaction between sleep disorders and processes, such as the glymphatic system, which has a circadian rhythm and increases during sleep, offering the opportunity to investigate the link between neurodegenerative diseases and sleep disorders, such as REM sleep behavior disorder or narcolepsy (29; 70).
The growing neuroimaging literature shows that insomnia is a multifaceted disorder that exhibits differences in brain function that are subtle and region-specific as well as state-specific (sleep vs. wakefulness). Therefore, neuroimaging can help in understanding subtypes of insomnia by better characterizing this heterogenous population using multiple metrics. Individuals with insomnia might have different structural and functional characteristics depending on which process is more affected.
Finally, neuroimaging can help assess the effect of a treatment intervention. For example, the effect of the first-line treatment for chronic insomnia, cognitive behavioral therapy for insomnia, on brain activity can be assessed with neuroimaging. The effect of continuous positive airway pressure treatment has been assessed in individuals with obstructive sleep apnea and shows a partial normalization of both symptoms and structural alterations in the hippocampus (12).
All contributors' financial relationships have been reviewed and mitigated to ensure that this and every other article is free from commercial bias.

Florence Pomares PhD
Dr. Pomares of Centre de Recherche de l'IUGM in Montreal has no relevant financial relationships to disclose.
See Profile
Nathan Cross PhD
Dr. Cross of Concordia University has no relevant financial relationships to disclose.
See Profile
Aurore A Perrault PhD
Dr. Perrault of Concordia University has no relevant financial relationships to disclose.
See Profile
Thien Thanh Dang-Vu MD PhD FAASM
Dr. Dang-Vu of Concordia University and CIUSSS du Centre-Sud-de-l’île-de-Montréal received speaker fees from Eisai, consultant fees from Idorsia, and research grants from Jass Pharmaceuticals and Paladin Labs as principal investigator.
See Profile
Antonio Culebras MD FAAN FAHA FAASM
Dr. Culebras of SUNY Upstate Medical University at Syracuse has no relevant financial relationships to disclose.
See ProfileNearly 3,000 illustrations, including video clips of neurologic disorders.
Every article is reviewed by our esteemed Editorial Board for accuracy and currency.
Full spectrum of neurology in 1,200 comprehensive articles.
Listen to MedLink on the go with Audio versions of each article.
MedLink, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Sleep Disorders
Jul. 05, 2026
Sleep Disorders
Jul. 03, 2026
Neurobehavioral & Cognitive Disorders
Jun. 17, 2026
Neuro-Oncology
May. 27, 2026
Neuropharmacology & Neurotherapeutics
May. 14, 2026
General Neurology
May. 13, 2026
Neuro-Oncology
Apr. 30, 2026
Neuropharmacology & Neurotherapeutics
Apr. 23, 2026