























Neurobehavioral & Cognitive Disorders
Mental status examination
Jun. 17, 2026
MedLink, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Worddefinition
At vero eos et accusamus et iusto odio dignissimos ducimus qui blanditiis praesentium voluptatum deleniti atque corrupti quos dolores et quas.
|
• Shellfish poisoning includes three neurotoxic syndromes that share some common features and are primarily associated with bivalve mollusks (eg, mussels, clams, oysters, and scallops). | |
|
• The neurologic shellfish poisoning syndromes are (1) amnesic shellfish poisoning, (2) neurotoxic shellfish poisoning, and (3) paralytic shellfish poisoning. | |
|
• About 40% of affected individuals require hospitalization, but fatalities are rare. | |
|
• Treatment is generally supportive and symptomatic. | |
|
• There are no available antitoxins for any of the neurologic forms of shellfish poisoning. | |
|
• Some survivors may have permanent neurologic disabilities after neurologic forms of shellfish poisoning, including potentially permanent short-term memory loss after amnesic shellfish poisoning. |
Toxic algal blooms and “red tides” have been documented for millennia (22). Ancient Greeks named the Red Sea for its appearance during red tides. The Bible refers to toxic algal blooms: “The waters that were in the rivers were turned to blood, and the fish in the rivers died, and the river stank...” (Exodus 7:20-21).
Shellfish poisoning includes four syndromes that share some common features and are primarily associated with bivalve mollusks (eg, mussels, clams, oysters, and scallops). As filter feeders, these shellfish can accumulate toxins produced by microscopic algae, such as cyanobacteria, diatoms, and dinoflagellates.
The neurologic shellfish poisoning syndromes are:
|
(1) Amnesic shellfish poisoning |
The fourth type of shellfish poisoning is diarrheal shellfish poisoning, which is outside of the scope of this article except as it concerns the differential diagnosis. Diarrheal shellfish poisoning toxins typically include okadaic acid and dinophysistoxins and can be considered to include the more recently identified toxin azaspiracid, although human disease due to the latter toxin is often separated as azaspiracid shellfish poisoning. These are considered briefly in tabular form under differential diagnosis.
Amnesic shellfish poisoning was first discovered in autumn 1987 when a serious outbreak of food poisoning occurred in eastern Canada (08; 79; 22). Three deaths and 105 confirmed cases of acute intoxication followed consumption of mussels from this area. Some surviving victims suffered long-term neurologic problems, including memory loss. The outbreak was traced to cultured blue mussels (Mytilus edulis) from the Cardigan Bay region of eastern Prince Edward Island. The toxin, identified as domoic acid, had not previously been found in any shellfish, and this outbreak was the first known occurrence of human poisoning by this neurotoxin. A plankton bloom at the time of the outbreak consisted almost entirely of the diatom Nitzschia pungens f. multiseries. Symptoms of intoxication in humans included abdominal cramps, vomiting within the first few hours, and neurologic abnormalities involving disorientation and memory loss that could persist indefinitely (25). In mid-December 1987, the toxin in mussels was identified as domoic acid (10; 25; 88).
The first paralytic shellfish poisoning event was reported in 1927 near San Francisco (56; 75), although clinical descriptions suggestive of paralytic shellfish poisoning have been documented for centuries (22). The outbreak was traced to shellfish contaminated with a toxin from the dinoflagellate Alexandrium catenella. There were six deaths among 102 individuals affected. This event eventually led to the institution of extensive shellfish monitoring programs in the United States and the establishment of regulatory limits on the associated neurotoxins in shellfish (66; 86).
|
• Individuals affected by amnesic shellfish poisoning typically report gastrointestinal symptoms (eg, vomiting, nausea, diarrhea, abdominal cramps, and hemorrhagic gastritis) from several hours to 3 days after ingestion of affected mollusks. In more severe cases, neurologic symptoms develop within 48 hours and include headache, dizziness, disorientation, visual disturbances, short-term memory loss, weakness, seizures, hiccups, and coma. | |
|
• The clinical presentation of neurotoxic shellfish poisoning typically includes both neurologic and gastrointestinal symptoms. | |
|
• The most common neurologic symptoms of neurotoxic shellfish poisoning include (1) numbness and tingling in the lips, mouth, face, and extremities; (2) dysesthesias (“nerves being on fire”); and (3) formication (sensation of “ants crawling and biting all over”). | |
|
• Symptoms of paralytic shellfish poisoning include headache, nausea and vomiting, dizziness, perioral and acral paresthesias, numbness (involving lips, mouth, face, neck, and extremities), limb weakness, dysarthria, respiratory failure, and, in severe cases, death. |
Amnesic shellfish poisoning. Affected individuals typically report gastrointestinal symptoms (eg, vomiting, nausea, diarrhea, abdominal cramps, and hemorrhagic gastritis) from several hours to three days after ingestion of affected mollusks. In more severe cases, neurologic symptoms develop within 48 hours and include headache, dizziness, disorientation, visual disturbances, short-term memory loss, weakness, seizures, hiccups, and coma. Other potential manifestations include profuse respiratory secretions, unstable blood pressure, and cardiac arrhythmias. Sequelae can include permanent short-term memory loss and peripheral polyneuropathy. Fatalities occur with very high doses of the toxin, often superimposed on comorbid conditions (eg, old age and renal failure).
Neurotoxic shellfish poisoning. Diagnosis of neurotoxic shellfish poisoning is based on the typical clinical presentation with a history of recent shellfish consumption or the development of respiratory symptoms after inhalation of toxins during red tide events (51). Symptoms typically begin 3 to 4 hours after toxin consumption but can begin anywhere from several minutes to 18 hours afterward (85). The clinical presentation typically includes both neurologic and gastrointestinal symptoms. The most common gastrointestinal symptoms are nausea, vomiting, and diarrhea. The most common neurologic symptoms are primarily neuropathic, including (1) numbness and tingling (paresthesia) in the lips, mouth, face, and extremities; (2) dysesthesias (“nerves being on fire”); and (3) formication (sensation of “ants crawling and biting all over”) (85). Less common neurologic symptoms can include ataxia, paresis or paralysis, reversal of hot and cold sensations, dysarthria, headache, mydriasis, and generalized fatigue (85).
Paralytic shellfish poisoning. Symptoms of paralytic shellfish poisoning include headache, nausea and vomiting, dizziness, perioral and acral paresthesias, numbness (involving lips, mouth, face, neck, and extremities), limb weakness, dysarthria, respiratory failure, and in severe cases, death. Ataxia has been reported but is not a common feature (40). There is no loss of consciousness, and the reflexes are unaltered. As opposed to tetrodotoxin poisoning (from ingestion of certain pufferfish), there is rarely significant hypotension. Gastrointestinal symptoms may be more prominent in Atlantic cases than in Pacific cases (34).
The typical course of intoxication with paralytic shellfish poisoning is fairly precipitous (13; 45). Symptoms start quickly: the median time between ingestion and onset is one hour, with a typical range between 30 minutes and 3 hours, although in some outbreaks the median incubation period was as long as 11 hours (20). Progression and intensity of symptoms vary with the amount of toxin ingested and intrinsic factors (eg, body weight, comorbidities, age). Children may be more sensitive to paralytic shellfish poisoning toxins than adults (69).
Five to 30 minutes following consumption of contaminated mollusks, there is mild perioral tingling progressing to numbness, which spreads to the face and neck in moderately affected cases. In severe cases, these symptoms spread to the extremities with incoordination and respiratory difficulty. Severe cases develop signs of medullary dysfunction with dysarthria, a sense of throat constriction, and incoherent speech or anarthria. In very severe cases and absence of ventilatory support, complete paralysis and death from respiratory failure can occur within 2 to 12 hours. After 12 hours, survivors gradually begin to recover and generally are asymptomatic within a few days (13; 35; 34).
In an outbreak affecting 31 people in the Philippines in 2013, all of whom consumed green mussel broth, the incubation period ranged from 1 to 23 hours (median 11 hours) (20). Most experienced neurologic symptoms: circumoral and extremity numbness (90%), dizziness (65%), and lightheadedness (55%). Eight cases (26%) were hospitalized, and two died (case fatality = 6%).
Tetrodotoxin shellfish poisoning. Tetrodotoxin shellfish poisoning is much less common and much less often recognized than other forms of shellfish poisoning. Tetrodotoxin has been documented in shellfish from Taiwan (19; 41), Japan (47; 60), Sicily (26), France (37), Spain (48), and Britain (82; 83; 28).
Human tetrodotoxin shellfish poisoning cases have been reported from Japan (47) and Taiwan (89). In particular, a significant outbreak following ingestion of gastropod molluscs occurred in Taiwan in 1994 involving 17 people (89). The molluscs consumed (ie, Nassarius castus and Nassarius conoidalis) contained various amounts of tetrodotoxin. Clinical features in these 17 cases resembled those conventionally found in tetrodotoxin poisoning, except that there was hypertension in eight cases (47%); the observed hypertension may have resulted from exaggerated responses to sympathetic stimuli or responses of the vasomotor center to a small dose of tetrodotoxin. All except one case had mild poisoning and recovered well.
The index case was a 71-year-old housewife with a history of hypertension and diabetes mellitus (89). In May 1994, 4 hours after eating more than 200 gastropod mollusks, she was brought to the emergency department at a local hospital with vomiting, general numbness, and weakness. She had bought these snails from her neighbor and had served them as a snack after her dinner. On arrival at the emergency department, she appeared drowsy and showed general malaise; her initial blood pressure was 210/100 mmHg, with a heart rate of 120 beats/minute. There was no evidence of respiratory distress, and routine physical examination did not reveal any other abnormal findings except slightly dilated pupils with prompt light reflex. Her blood pressure normalized by the next morning, but her general weakness progressed gradually, and she became cyanotic. Endotracheal intubation, as well as assisted ventilatory support, were then performed, and she later regained consciousness. She removed her endotracheal tube 1 day later but was reintubated due to massive aspiration pneumonia and obvious respiratory distress. Her aspiration pneumonia resolved slowly with parenteral antibiotics and mechanical ventilation. Acute renal failure developed 1 week later, followed by upper gastrointestinal bleeding. Although she was treated with regular dialysis and blood component therapy, her condition deteriorated, and 27 days after her admission, she died from multiple organ failure.
Memory loss associated with amnesic shellfish poisoning may persist indefinitely (25).
Fatalities following amnesic shellfish poisoning were reported with the initial outbreak in eastern Canada in 1987: specifically, three deaths among 105 confirmed cases of acute intoxication followed consumption of mussels from this area.
Since 1995, there have been multiple outbreaks in the United States with 114 affected individuals, about 40% of whom required hospitalization, but no deaths.
An 84-year-old, previously healthy man developed nausea and vomiting after eating mussels (15). The following day, he became progressively disoriented and somnolent, and by the third day, he was comatose and had complex partial status epilepticus. His seizures did not respond to intravenous phenytoin, and large doses of phenobarbital were required to control them. EEGs showed diffuse slowing of background activity, periodic lateralized epileptiform discharges (PLEDs) over the left hemisphere, and later bitemporal independent epileptic abnormalities. No abnormality was observed on head CT scans with and without contrast, except for mild ventricular enlargement and cerebral atrophy consistent with his age. After several weeks, he slowly began to improve. After more than four months, he was discharged from the hospital on phenobarbital and was seizure-free but had severely impaired anterograde memory.
Mussels from the same area as those he had eaten were later shown to contain domoic acid.
Phenobarbital was discontinued after two months, and an EEG obtained eight months after the intoxication was normal. However, one year after the acute episode, he developed complex partial seizures with secondary generalization. Seizures did not recur after he received intravenous phenytoin. EEGs showed epileptic discharges over both temporal lobes. On MRI, T2-weighted images revealed hyperintense signal and marked atrophy of both hippocampi. A PET scan showed a bitemporal decrease in glucose metabolism. Neuropsychological evaluation revealed a normal IQ and normal cognition, but his verbal and nonverbal memory was severely impaired. He was discharged on phenytoin 300 mg daily, with good seizure control thereafter.
Three and a half years after intoxication, he developed pneumonia and died. At autopsy, there was obvious atrophy of the hippocampi and slight dilatation of the ventricular system and of the Sylvian fissure. The hippocampi showed complete neuronal loss in CAI and CA3, almost total loss in CA4, and moderate loss in CA2.
|
• The frequency of harmful algal blooms has been increasing in recent decades due to human activities, climate change, and the eutrophication of marine waters. | |
|
• When toxins from the algal blooms are accumulated in high concentrations by shellfish during filter feeding of plankton, the toxins can be passed on to birds, marine mammals, and humans by consumption of the contaminated shellfish. | |
|
• Each of the neurologic forms of shellfish poisoning is caused by a specific toxin produced by planktonic diatoms (group of single-celled microalgae with cell walls of silica): (1) amnesic shellfish poisoning is caused by consumption of domoic acid; (2) neurotoxic shellfish poisoning is caused by consumption of brevetoxins; (3) paralytic shellfish poisoning is caused by consumption of saxitoxin. |
The frequency of harmful algal blooms along coastlines has been increasing in recent decades due to human activities, climate change, and the eutrophication of marine waters (ie, over-abundance of plant nutrients from agricultural runoff, deforestation, and erosion) (90). A major contributor to coastal algal blooms is the release of excess nutrients (eg, nitrogen or phosphorus) from fertilizer runoff entering the aquatic system and causing excessive growth of algae. Harmful algal blooms commonly cause the widespread death of fish and seabirds.
Dinoflagellate blooms are commonly called “red tides,” but they can occur with other color changes in water (eg, green, brown, or yellow), and toxic levels of the associated toxins can occur in clear-appearing water. Consequently, such events should be called “harmful algal blooms” instead of “red tides” (74; 43; 40).
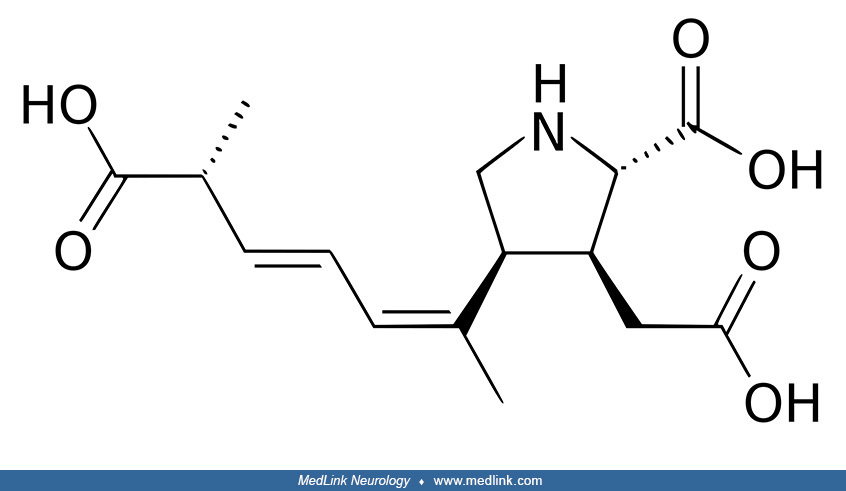
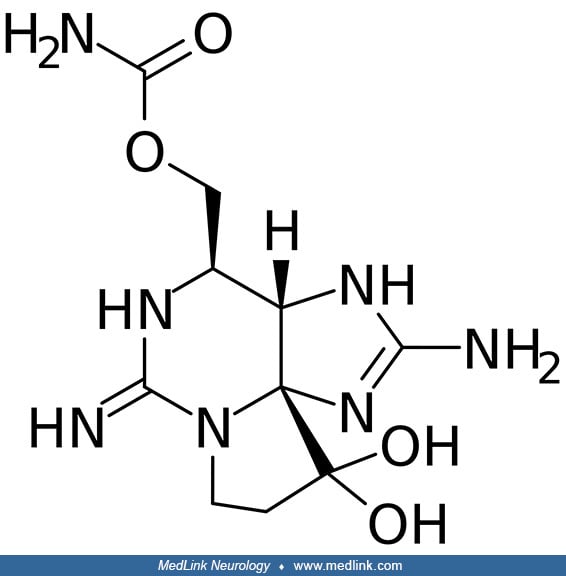
Amnesic shellfish poisoning. Amnesic shellfish poisoning is an illness caused by consumption of the marine biotoxin called domoic acid, which structurally resembles both the excitatory neurotransmitter glutamate and the neurotoxin kainic acid (79; 42; 33; 62).
In mammals, including humans, domoic acid acts as a neurotoxin, potentially causing permanent short-term memory loss and, in severe cases, death.
Domoic acid was first isolated in 1959 from a species of red algae (Chondria armata) in Japan. Domoic acid is produced naturally by marine diatoms belonging to the genus Pseudo-nitzschia and the species Nitzschia navis-varingica. Some fishery resources of high commercial value, such as the king scallop Pecten maximus, are frequently exposed to toxic Pseudo-nitzschia blooms and are capable of accumulating high amounts of domoic acid, retaining it for months or even a few years (30). Diatoms are a major group of algae, specifically single-celled microalgae with a cell wall of silica.
In the brain, domoic acid preferentially damages the hippocampus and amygdaloid nucleus by activating AMPA (alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) and kainate receptors (a class of ionotropic receptors that respond to the neurotransmitter glutamate), causing an uncontrolled influx of calcium (25; 15; 22; 23; 36; 91; 70). Uncontrolled increase of calcium ions causes affected cells to degenerate (67; 91). Domoic acid is also an excitatory amino acid analogue of glutamate with a very strong affinity for glutamate receptors, which results in excitotoxicity, compounded by a synergistic effect with endogenous glutamate and N-methyl-D- aspartate receptor agonists (15). Seizures following acute domoic acid intoxication and the postmortem pathology of the hippocampus (with profound loss of pyramidal cells) indicate that (1) the hippocampus is vulnerable to kainate receptor excitotoxicity, and (2) kainate-receptor-induced excitotoxic injury may play a role in epileptogenesis (15).
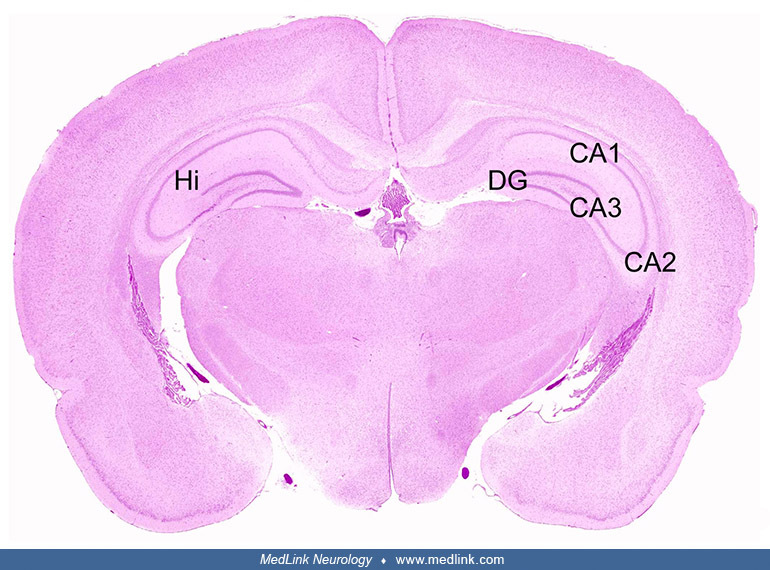
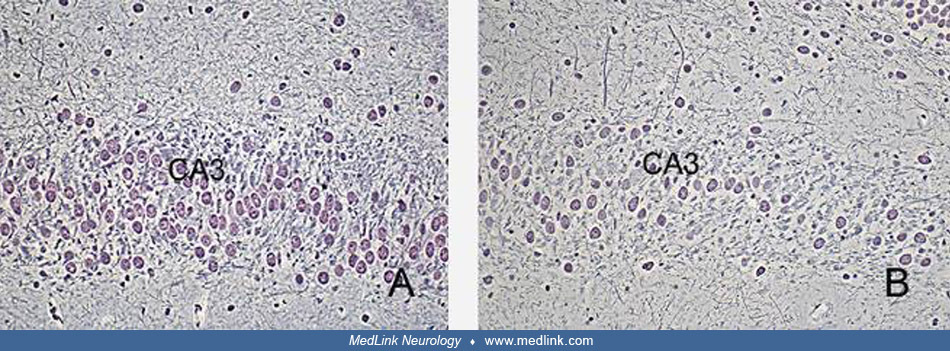
The neurotoxic effects of domoic acid have been demonstrated in many mammalian species (67). In rats exposed to domoic acid, there is cell dropout and necrosis, particularly within the CA3 regions of the hippocampus, as shown at different magnifications in sections stained with hematoxylin and eosin (67).
Cross-section of both hippocampal formations (Hi) showing the dentate gyrus (DG), and the CA3, CA2, and CA1 regions. Hematoxylin and eosin staining. (Source: Pulido 2008. Creative Commons Attribution license. http://creativecom...
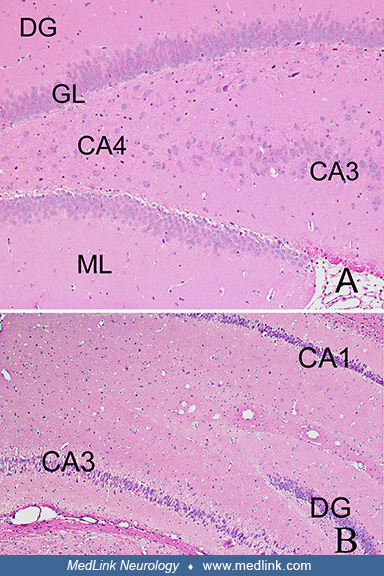
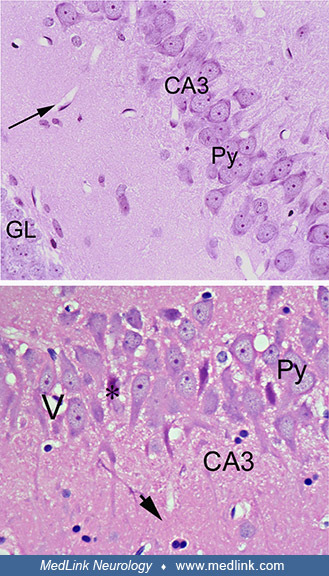
Top image (control rat) is a cross-section of the hippocampus of a control rat, showing the granular cell layer (GL) of the dentate gyrus (DG), the molecular layer (ML), and the CA3 and the CA4 regions. Objective x10. Bottom im...
Top image (control rat) shows the CA3 region with well-preserved pyramidal cells (Py). Blood vessels are seen as white spaces with the endothelial cells at the periphery (arrow). Objective x40. Bottom image (domoic-acid-treated...
At higher magnifications, vacuolar cytoplasm within pyramidal neurons in the CA3 region is evident. Silver stains show a marked dropout of pyramidal neurons in the CA3 region (67).
(Left) Hippocampus of a control rat. CA3 regions. Chen/Bodian silver stain, Objective x20. (Right) Sections of the hippocampus of a rat treated with domoic acid, showing-marked drop out of pyramidal neurons in the CA3 region. C...
Monkeys exposed to domoic acid also show cell dropout and neuronal necrosis in the CA3 region of the hippocampus in sections stained with hematoxylin and eosin (67).
Brains of monkeys (Macaca fascicularis, the crab-eating macaque). (Left) Section of the CA3 region of the hippocampus of a control monkey, showing well-preserved pyramidal cells (Py). Blood vessels are seen as white sp...
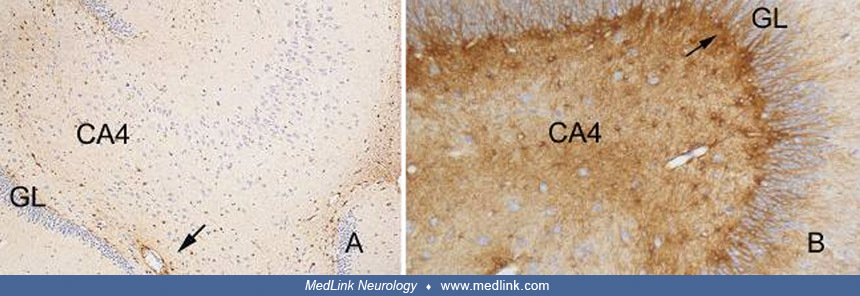
Most pyramidal neurons appear with vacuolar cytoplasm, and some nuclear pyknosis is also evident. Immunohistochemical staining of monkey brains for glial fibrillary acidic protein (GFAP) shows marked astrocytosis as revealed by the intensity of the staining seen in CA4 and the subgranular zone (67).
Brains of monkeys (Macaca fascicularis, the crab-eating macaque). Histological sections were processed for GFAP immunohistochemistry. (Left) Hippocampus of a control monkey showing the granular cell layer (GL) of the d...
The retina is also affected by domoic acid neurotoxicity, as shown in the stained eye of a monkey treated with a single intravenous dose of domoic acid; there is cell loss and necrosis, particularly in the inner nuclear and ganglion cell layers (67).
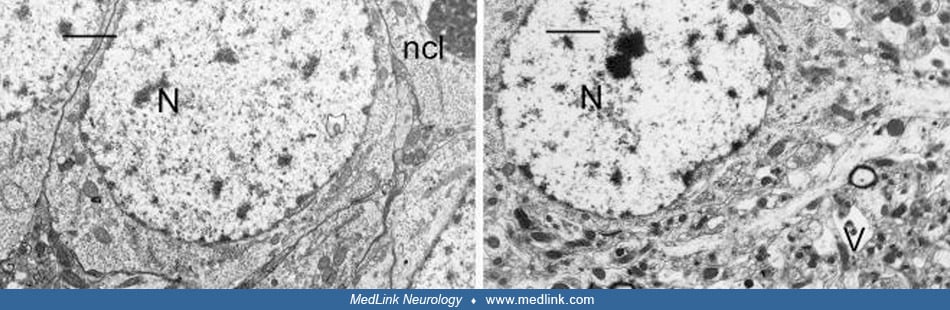
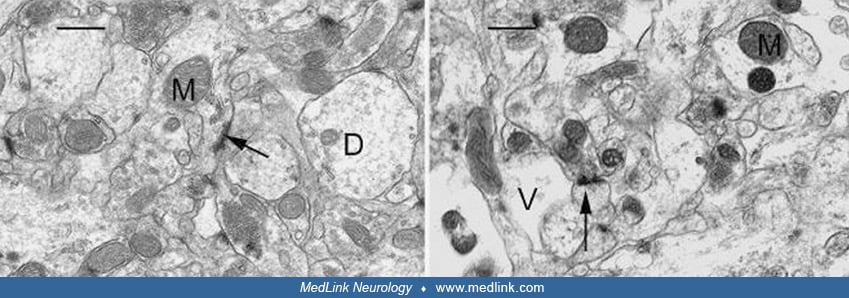
Electron microscopy of rats treated with domoic acid shows vacuolization of pyramidal neurons in the CA3 region (67).
(Left) CA3 region of the hippocampus of a control rat showing a cluster of pyramidal cells with good preservation and integrity of cell membranes and organelles. The nucleus (N) and nucleolus (ncl) are easily identified. A well...
(Left) CA3 region of the hippocampus of a control rat showing good preservation and integrity of the neuropil. Mitochondria (M), synaptic spines (arrow) and dendrites (D) are identified. Scale Bar = x1.1μm. (Right) CA3 region o...
(Left) High magnification of the CA3 region of the hippocampus of a control rat showing good preservation and structural integrity. An electrodense dendritic spine (arrow), terminal axon (At), dendrite, and mitochondria (M) are...
The presence of numerous vacuoles of various sizes in pyramidal cells has been referred to as the “Swiss cheese” effect for its obvious similarity to the profusion of round holes (“eyes”) in this type of cheese. At higher magnification, the loss of structural integrity and marked vacuolar dilatation of dendrites are evident (67).
Long-term treatment of primary mesencephalic cell cultures with domoic acid destroys dopaminergic neurons (68). Domoic acid significantly decreased the number of dopaminergic neurons and adversely affected their morphology. Co-treatment of cultures with domoic acid and AMPA/KA or NMDA receptor antagonists rescued a significant number of dopaminergic neurons.
Climate change is projected to cause considerable contraction and potential extirpation for all three key amnesic shellfish toxin (AST)-producing diatom species (ie, Pseudo-nitzschia australis, P seriata, and P fraudulenta) at lower latitudes together with projected poleward expansions into higher latitudes, mainly in the northern hemisphere (12).
The observation of the neurotoxin domoic acid in different geographic regions and water masses of Alaska across various Pseudo-nitzschia species Fragilariopsis oceanica, and in different seasons, underscores the significant risk of amnesic shellfish poisoning and neurotoxin domoic acid poisoning in Alaska waters (38).
Neurotoxic shellfish poisoning. Neurotoxic shellfish poisoning is caused by consumption of brevetoxins, which are marine toxins produced primarily by the dinoflagellate Karenia brevis (previously known as Gymnodinium breve and Ptychodiscus brevis), although several other dinoflagellates may also produce these toxins (42; 02; 33).
These toxins can produce a series of gastrointestinal and neurologic effects. Outbreaks of neurotoxic shellfish poisoning commonly occur following harmful algal bloom events, which are more commonplace along the coasts of Florida and Texas, especially during late summer and early fall.
Human exposure most commonly occurs from the consumption of commonly harvested shellfish, such as clams, oysters, and mussels. However, exposure to lower levels of brevetoxins can occur with consumption of certain planktivorous fish (85). Toxins are present in the flesh of shellfish for 2 to 8 weeks following a harmful algal bloom, although in some cases, toxin may be retained for nearly one year post-bloom (85).
Brevetoxins are a group of more than 10 lipid-soluble cyclic polyethers that bind to a specific site on the voltage-gated sodium channel, leading to an influx of sodium ions into the cell (65; 80; 49). This results in activation of nerves and spontaneous nerve cell membrane depolarization and firing (84; 85). Additionally, brevetoxins can bind a separate site on voltage-gated sodium channels, causing release of neurotransmitters (such as acetylcholine), resulting in tracheal smooth muscle contraction and widespread mast cell degranulation (84). Because of their lipid-solubility, brevetoxins pass through cell membranes and cross the blood-brain barrier (85). They are metabolized primarily by the liver and are excreted in the bile, although urinary excretion also aids toxin clearance (85). Brevetoxins can be highly concentrated in the sea spray aerosol generated by waves hitting the shore during a red tide event (63; 64).
Paralytic shellfish poisoning. Like the other neurotoxic forms of shellfish poisoning, paralytic shellfish poisoning is primarily associated with bivalve mollusks (eg, mussels, clams, oysters, and scallops). These shellfish are filter feeders and accumulate the neurotoxin saxitoxin, produced by microscopic algae, such as dinoflagellates, diatoms, and cyanobacteria (22; 42; 29; 33). Dinoflagellates of the genus Alexandrium are the most numerous and widespread producers of paralytic shellfish toxins.
Paralytic shellfish toxins, which are widely distributed in marine environments, are a group of neurotoxic alkaloids, more than 60 of which have been identified to date (90).
Saxitoxin is the most potent of the paralytic shellfish toxins (42).
Saxitoxin, is similar to tetrodotoxin but much more potent (44; 06; 42). Saxitoxin and its derivatives, including gonyautoxins, are water-soluble, heat-stable, tetrahydropurine compounds (87; 11; 71; 72; 29; 24).
These neurotoxins block tetrodotoxin-sensitive voltage-gated Na+ channels, preventing entry of Na+ into cells; this disrupts nerve conduction in motor and sensory nerves (29; 24; 27).
(A) Structural organization of the voltage-gated sodium channel complex. The voltage-gated sodium channel complex is composed of a pore-forming alpha subunit and one or more beta subunits. The alpha subunit, a single polypeptid...
Saxitoxin produces and is responsible for paralytic shellfish poisoning blooms in subarctic(including Alaska), temperate(including Oregon), and tropical locations (07; 14; 76; 42; 50; 78; 81). In Asia, paralytic shellfish poisoning is mostly associated with the species Pyrodinium bahamense (05). Human intoxication usually occurs through the consumption of paralytic toxin-contaminated bivalve shellfish (ie, mussels, oysters, and clams), although some pufferfish also contain saxitoxin, making their consumption hazardous (42; 59).
Alexandrium catenella has been a significant threat to human health in southeastern Alaska for centuries (03). It is transported into Arctic regions in water moving northward through the Bering Strait. An enormous A catenella benthic cyst bed and hydrographic conditions across the Chukchi Sea support germination and development of recurrent, locally originating, and self-seeding blooms. Over the past two decades, warming has increased the magnitude of the germination flux twofold and advanced the timing of cell inoculation into the euphotic zone by 20 days, making conditions favorable for bloom development in surface waters. Such annually recurrent and massive A catenella blooms pose a significant threat to public and ecosystem health in Alaskan Arctic communities.
Similarly, in British Columbia, Canada, documented poisonings are increasing in frequency, coincident with rising global temperatures, based on 62 investigations identified from 1941 to 2020 (54). Most paralytic shellfish poisoning investigations (71%) and cases (69%) were linked to self-harvested shellfish, particularly by indigenous communities. Single-case exposure investigations increased by more than 3.5 times in the decade 2011 to 2020 compared to previous periods. Clams (47%), mussels (26%), oysters (14%), scallops (6%), and, in more recent years, crabs (4%) were linked to illnesses.
A catenella also shows an apparent oceanic range expansion in the southern hemisphere from south to the north since its first detection in 1972 (61).
Tetrodotoxin shellfish poisoning. Tetrodotoxin (TTX), a potent neurotoxin mostly associated with pufferfish poisoning, is also found in bivalve shellfish.
Tetrodotoxin is a sodium channel blocker that inhibits action potentials in neurons by binding to the voltage-gated sodium channels in nerve cell membranes and blocking the passage of sodium ions (responsible for the rising phase of an action potential) into the neuron.
Tetrodotoxin has been recognized in shellfish in Japan and Taiwan, and in a few, mainly estuarine shellfish production areas in European countries. Both tetrodotoxin and paralytic shellfish toxins can be detected in organisms that possess either of them (46).
Water temperature appears to be one of the key triggers of events leading to tetrodotoxin accumulation in European bivalves, although other factors are also likely important (eg, the presence of a de novo biological source). A large systematic tetrodotoxin screening study, encompassing over 3500 bivalve samples collected throughout 2016 from 155 shellfish monitoring sites along the coast of Great Britain, found that only 1% of tested samples contained tetrodotoxin above the reporting limit of 2 μg/kg whole shellfish flesh; these samples all originated from ten shellfish production sites in southern England (28). Subsequent continuous monitoring of selected areas over a 5-year period showed a seasonal tetrodotoxin accumulation in bivalves, starting in June when water temperatures reached around 15 °C.
Tetrodotoxin (TTX) concentrations in bivalves and sea-surface temperature (SST) over multiple years. The data were collected from selected shellfish representative monitoring sites in the United Kingdom. (Source: Dhanji-Rapkova...
Sea-surface temperature characteristics of selected shellfish representative tetrodotoxin (TTX) monitoring sites in the United Kigndom where TTX-positive or TTX-negative bivalves were recovered in 2016. Box and whisker plots sh...
Daily continuous sea-surface temperature (SST) changes at selected shellfish representative tetrodotoxin (TTX) monitoring sites in the United Kingdom where TTX-positive or TTX-negative bivalves were recovered in 2016. Satellite...
Magnified view of period corresponding to tetrodotoxin (TTX) accumulation in bivalves at selected shellfish representative monitoring sites in 2016. The plot shows satellite-derived daily mean sea-surface temperature (SST) valu...
|
• Since 1995, there have been 28 recorded outbreaks of neurologic forms of shellfish poisoning in the United States. | |
|
• Since 1995, among 114 individuals who were sickened with neurologic forms of shellfish poisoning in the United States, about 40% required hospitalization, but none were fatal. |
Toxic algal blooms have potentially serious consequences for human and animal populations. In some cases, monitoring “red tide” events, sampling shellfish beds, and observing effects on local wildlife can identify conditions when shellfish poisoning is likely. For example, an outbreak of shellfish poisoning in England in 1968 affected 68 people (53); this, however, was preceded by a profuse multiplication of dinoflagellates off the Northumbrian coast, followed by finding dead sand-eels that probably ingested the toxin by feeding on marine fauna that subsist on dinoflagellates, and then birds were seriously affected by eating the dead and dying sand-eels (21). In this case, with public notifications via television and radio, effective control measures were instituted quickly and proved to be effective (53). Testing of shellfish over a wide expanse of the coastline made it possible to define the area of risk and to target preventive measures.
Reports of harmful algal blooms in United States waters and around the world have markedly increased over the past four decades. The increased risks to humans of shellfish toxicity from harmful algal blooms is a consequence of large-scale ecological changes from anthropogenic activities, especially increased eutrophication, marine transport and aquaculture, and global climate change (43; 03; 54; 55; 61).
A 30-year review of harmful algal events in Canada from 1988 to 2017 documented that recurring harmful algal events have been widespread throughout both the Atlantic and Pacific coastal regions of Canada (55). The 367 paralytic shellfish toxin reports revealed annual and frequent recurrence throughout Canada’s Atlantic and Pacific regions, including multiyear paralytic shellfish poisoning events in the Bay of Fundy, the Estuary and Gulf of St. Lawrence, and the Strait of Georgia.
In the United States, nearly half of foodborne disease outbreaks of chemical origin are caused by toxic fish or shellfish (39).
The U.S. National Office for Harmful Algal Blooms defines harmful algal bloom events as situations that constituted a quarantine, closure, or management decision where toxicity was detected exceeding regulatory limits.
Amnesic shellfish poisoning. The first outbreak of amnesic shellfish poisoning was recorded in 1987. There were three deaths among 105 confirmed cases in eastern Canada around Prince Edward Island (08; 79; 22; 55).
Data compiled by the U.S. National Office for Harmful Algal Blooms in the period from 2011 to 2020 show that presence of the toxin for amnestic shellfish poisoning (ie, domoic acid) exceeded regulatory limits frequently along the west coast and the northeast, with some events along the Gulf Coast of Florida and the coast of Alaska. Internationally, the greatest concentration of amnestic shellfish poisoning occurs in North America (especially Canada and the United States), Europe, Australia, and New Zealand, with some events also noted in Chile.
According to data from the National Outbreak Reporting System (NORS) of the Centers for Disease Control and Prevention, the only recognized outbreak of human disease in the United States occurred in New Jersey in July 2013, when two individuals were hospitalized with amnesic shellfish poisoning after eating mollusks at a restaurant.
When accumulated in high concentrations by shellfish during filter feeding of plankton, domoic acid can be passed on to birds, marine mammals, and humans by consumption of the contaminated shellfish. Human illness due to domoic acid has only been recognized following shellfish consumption (15), even though the toxin can bioaccumulate in many marine organisms that consume phytoplankton, such as anchovies and sardines.
In 2015, the Pacific coast of North America was heavily impacted by an algal bloom, consisting predominantly of the domoic acid-producing pennate diatom, Pseudo-nitzschia. Consequently, elevated levels of domoic acid were measured in stranded marine mammals, prompting the closure of beaches and damaging razor clam, rock crab, and Dungeness crab fisheries.
Neurotoxic shellfish poisoning. In October 1987, a red tide due to Karenia brevis (previously known as Gymnodinium breve and Ptychodiscus brevis) affected the North Carolina coast for the first time (57). Forty-eight people who had eaten oysters at 20 meals developed gastrointestinal and neurologic symptoms from October 27 to December 9. About half of the cases occurred after the first closure of the shellfish waters on November 2. There was a dose-response effect, with a significant increase in illness attack rate with increasing numbers of oysters eaten. The illnesses were generally mild and of short duration, with no deaths reported.
Data compiled by the U.S. National Office for Harmful Algal Blooms in the period from 2011 to 2020 show that the presence of the toxins for neurotoxic shellfish poisoning (ie, brevetoxins) exceeded regulatory limits frequently along the Gulf Coast, particularly along the coast of Florida. Internationally, the greatest concentration of neurotoxic shellfish poisoning occurs along the Atlantic and Gulf coasts of North America, in several lakes in northeast Germany, and along the coasts of New Zealand.
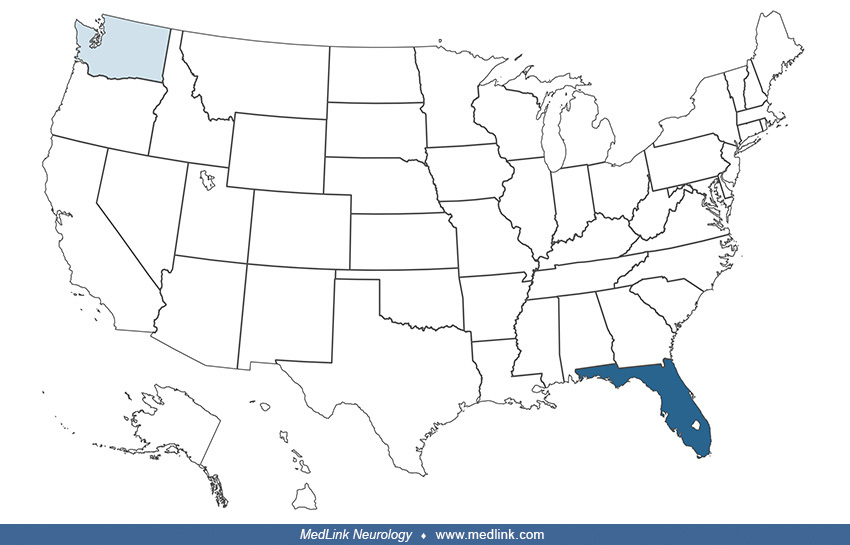
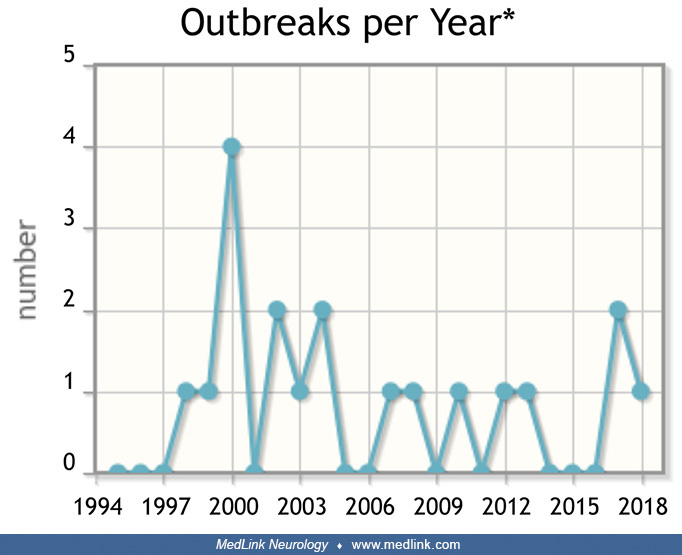
According to data from the National Outbreak Reporting System (NORS) of the Centers for Disease Control and Prevention, there have been eight outbreaks of neurotoxic shellfish poisoning since 1995 (Table 1); seven outbreaks were seen in Florida, and one in Washington. Collectively, there were 31 identified individuals clinically symptomatic, with 12 hospitalizations and no deaths. Neurotoxic shellfish poisoning may occur at any time of the year and does not appear to have a seasonal pattern.
|
Year |
Month |
State |
Etiology Status |
Setting |
Illnesses |
Hospitalizations |
Deaths |
Food eaten |
|
2000 |
6 |
WA |
Suspected |
Restaurant |
3 |
0 |
0 |
steamed mussels |
|
2001 |
3 |
FL |
Suspected |
Restaurant |
2 |
0 |
0 |
shrimp (unspecified) |
|
2001 |
11 |
FL |
Suspected |
Private residence |
2 |
1 |
0 |
seafood chowder |
|
2005 |
7 |
FL |
Confirmed |
Private residence |
4 |
3 |
0 |
steamed clams |
|
2006 |
7 |
FL |
Confirmed |
Private residence |
13 |
4 |
0 |
clams |
|
2006 |
10 |
FL |
Confirmed |
Private residence |
2 |
1 |
0 |
raw clams |
|
2007 |
1 |
FL |
Suspected |
Private residence |
3 |
0 |
0 |
steamed clams |
|
2017 |
3 |
FL |
Confirmed |
Private residence |
2 |
1 |
0 |
conch soup |
|
| ||||||||
In southwest Florida, a dose-response relationship was observed between K brevis blooms and reporting of upper respiratory tract and neurotoxic shellfish poisoning-like symptoms and headaches (01); reporting of neurotoxic shellfish poisoning-like symptoms was higher among participants with a history of migraines, chronic fatigue syndrome, and mild memory loss, whereas the association of headaches with K brevis blooms was accentuated among individuals with a history of migraines.
Paralytic shellfish poisoning. The case-fatality rate of paralytic shellfish poisoning was about 9% to 10% in some early large series (04).
An outbreak of paralytic shellfish poisoning occurred in Champerico, on the Pacific coast of Guatemala, in July and August of 1987 (69). Of 187 people affected, 26 died (14%). A species of clam, Amphichaena kindermani, harvested from local beaches, was the source of the neurotoxins (saxitoxins). Children younger than six years of age had a higher case-fatality rate (50%) than adults (with a case-fatality rate of 7% among those older than 18 years of age).
In Alaska from 1973 to 1992, 54 outbreaks of paralytic shellfish poisoning involved 117 people (31).
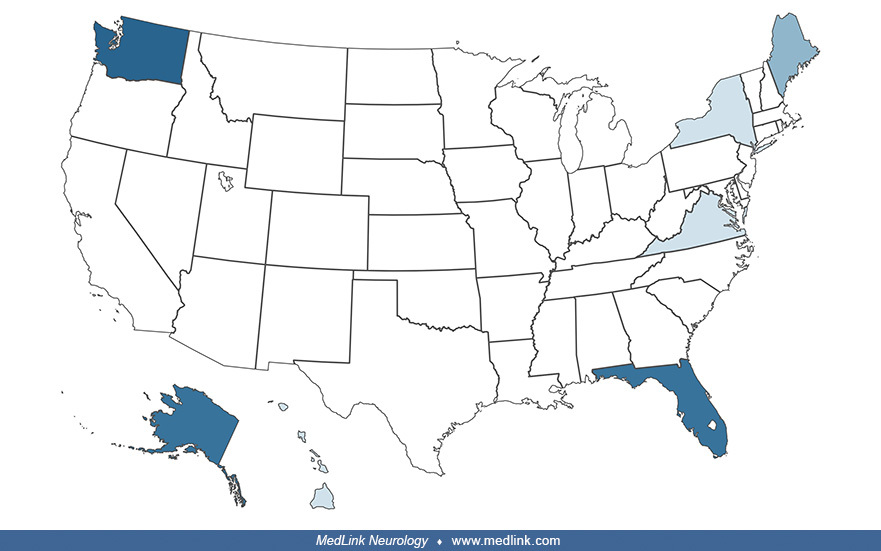
According to data from the National Outbreak Reporting System (NORS) of the Centers for Disease Control and Prevention, there have been 19 outbreaks of paralytic shellfish poisoning since 1995 (Table 2), most commonly involving Florida, Washington, and Alaska, but with smaller numbers involving Maine, New York, Virginia, and Hawaii. Collectively, there were 81 identified individuals clinically symptomatic, with 30 hospitalizations and no deaths. Paralytic shellfish poisoning tends to be more common in the summer months, but it may occur at any time of the year.
|
Year |
Month |
State |
Etiology Status |
Setting |
Illnesses |
Hospitalizations |
Deaths |
Food eaten |
|
1998 |
10 |
WA |
Confirmed |
3 |
2 |
0 |
clams | |
|
1999 |
3 |
HI |
Confirmed |
Private residence |
7 |
4 |
0 |
mussels |
|
2000 |
8 |
WA |
Confirmed |
2 |
0 |
0 |
clams | |
|
2000 |
8 |
WA |
Suspected |
3 |
papio | |||
|
2000 |
8 |
WA |
Confirmed |
Private residence |
2 |
mussels, unspecified | ||
|
2000 |
6 |
WA |
Confirmed |
Private residence |
3 |
3 |
mussels, unspecified | |
|
2002 |
9 |
AK |
Confirmed |
Private residence |
4 |
3 |
steamed mussels | |
|
2002 |
1 |
multiple |
Suspected |
Restaurant |
3 |
steamed mussels | ||
|
2003 |
4 |
FL |
Confirmed |
Other |
6 |
mussels, unspecified | ||
|
2004 |
2 |
FL |
Suspected |
Private residence |
4 |
1 |
0 |
mussels, unspecified |
|
2004 |
4 |
FL |
Confirmed |
Private residence |
2 |
2 |
0 |
pufferfish |
|
2007 |
7 |
ME |
Confirmed |
Private residence |
2 |
1 |
0 |
pufferfish |
|
2008 |
7 |
ME |
Confirmed |
Private residence |
2 |
1 |
0 |
pufferfish |
|
2010 |
6 |
AK |
Suspected |
Private residence |
21 |
6 |
0 |
pufferfish |
|
2012 |
9 |
WA |
Confirmed |
4 |
4 |
0 |
steamed mussels | |
|
2013 |
10 |
AK |
Confirmed |
3 |
3 |
0 |
steamed mussels | |
|
2017 |
7 |
AK |
Confirmed |
Private residence |
5 |
0 |
0 |
mussels, unspecified |
|
2017 |
7 |
AK |
Confirmed |
Private residence |
3 |
0 |
0 |
steamed clams |
|
2018 |
6 |
FL |
Suspected |
Private residence |
2 |
0 |
0 |
pufferfish |
|
Source: National Outbreak Reporting System (NORS) of the Centers for Disease Control and Prevention | ||||||||
The only means of preventing neurologic forms of shellfish poisoning is to refrain from eating potentially contaminated shellfish obtained from high-risk areas (ie, coastal areas during red tides).
Amnesic shellfish poisoning. Shellfish containing toxic levels of domoic acid do not look or taste any different from shellfish that are safe to eat. Laboratory testing of shellfish meat is the only known method of detecting the neurotoxin. Furthermore, domoic acid is a very stable, heat-resistant toxin; consequently, cooking or freezing affected fish or shellfish does not lessen the toxicity.
Razor clams are most often affected because the algae that produce domoic acid are more commonly found in coastal areas. Domoic acid has also been detected in mussels, clams, and oysters. Dungeness crabs, because they feed on razor clams and other shellfish, can also become toxic. Even if the crab meat is safe, domoic acid tends to accumulate in the crab gut and "butter" (ie, the white-yellow fat inside the back of the shell). Before cooking, crabs should be cleaned thoroughly by removing all the butter and discarding the guts.
Neurotoxic shellfish poisoning. Brevetoxins are tasteless and odorless and cannot be eradicated by rinsing, cleaning, cooking, freezing, or the application of acid (84; 85). There is presently no means of preventing the uptake of toxins by shellfish nor of removing the toxins from shellfish after harvest (58).
In the United States, coastal states take measures to monitor and prevent shellfish poisoning outbreaks. For example, the Florida Department of Health has added neurotoxic shellfish poisoning to its list of reportable diseases. In addition, the Florida Department of Environmental Protection monitors dinoflagellate levels and restricts the harvest of shellfish when levels of brevetoxin are dangerously elevated during Florida red tide events. Shellfish beds are subsequently opened after two weeks if mouse bioassay testing confirms safe levels (85).
Paralytic shellfish poisoning. According to the Centers for Disease Control and Prevention, avoidance of noncommercially harvested shellfish not tested for saxitoxins is the best way to prevent paralytic shellfish poisoning (16). In Alaska, for example, commercially harvested shellfish are tested for saxitoxin and considered safe for human consumption, but shellfish collected by persons for their own use are not tested and may be contaminated (16).
The differential diagnosis of an acute gastrointestinal illness with recent shellfish ingestion would include diarrheic shellfish poisoning and bacterial or viral gastroenteritis. With neurologic forms of shellfish poisoning, gastrointestinal symptoms may begin before neurologic symptoms.
The various neurologic forms of shellfish poisoning must be distinguished from one another, with the salient distinguishing features being short-term memory loss with amnesic shellfish poisoning, weakness with paralytic shellfish poisoning, and other neurotoxic symptoms with neurotoxic shellfish poisoning (Table 3). In addition, the neurologic forms of shellfish poisoning need to be distinguished from ciguatera, pufferfish poisoning, scombroid fish poisoning (from eating spoiled fish that have naturally high levels of histidine because the histidine is converted to histamine with spoilage), seafood allergy, organophosphate pesticide poisoning, alcohol intoxication, and certain psychiatric disorders (85).
|
|
Amnesic shellfish poisoning |
Neurotoxic shellfish poisoning |
Paralytic shellfish poisoning |
Diarrheic shellfish poisoning |
Azaspiracid shellfish poisoning |
Ciguatera fish poisoning |
|
Toxin-producing organism |
Diatoms: Pseudo-nitzchia species |
Dinoflagellates: Karenia brevis and other Karenia species |
Dinoflagellates: Alexandrium species, Gymonodinium catenatum, Pyrodinium bahamense |
Dinoflagellates: Dinophysis species, Prorocentrum lima |
Dinoflagellates: Proroperidiunium species |
Dinoflagellates: Gambierdiscus toxicus, possibly others |
|
Toxin(s) |
Domoic acid |
Brevetoxins |
Saxitoxins |
Okadaic acid |
Azaspiracid |
Ciguatoxins |
|
Foods likely to be contaminated |
Shellfish, primarily scallops, mussels, clams, oysters |
Shellfish, primarily mussels, oysters, scallops |
Shellfish, primarily scallops, mussels, clams, oysters, and cockles |
Shellfish, primarily scallops, mussels, clams, oysters |
Shellfish |
Reef fish, such as barracuda, grouper, red snapper, and amberjack |
|
Short-term symptoms |
Nausea |
Nausea |
Nausea |
Nausea |
Nausea |
Nausea |
|
Long-term symptoms |
Possibly amnesia |
Unknown |
Unknown |
Unknown |
Unknown |
Abnormal hot and cold sensations |
|
Treatment |
Supportive care, especially for older people and those with kidney disease |
Supportive care |
Supportive care |
Supportive care |
Supportive care |
Supportive care |
|
| ||||||
Many cases of neurologic shellfish poisoning have gone unrecognized. In a retrospective study of poisoning cases by bivalve shellfish (oysters, mussels, and scallops) recorded by the French Poison Control Centers from 2012 to 2019, 22% (n=134) of the 619 shellfish poisoning cases recorded had reported at least one neurologic symptom (eg, headache, dizziness, or paraesthesia) (73). A review of medical records for these 134 patients led to suspicion of 14 cases of paralytic shellfish poisoning and one case of amnesic shellfish poisoning.
Diagnosis of neurologic forms of shellfish poisoning is based on the typical clinical presentation with a history of recent shellfish consumption or the development of respiratory symptoms after inhalation of toxins during red tide events.
Some shellfish toxins can be detected in serum and urine specimens (32).
The Centers for Disease Control and Prevention (CDC) has established case definitions for brevetoxin and saxitoxin poisoning but not for domoic acid poisoning (17; 18).
According to the CDC case definition of brevetoxin poisoning, as occurs with neurotoxic shellfish poisoning, laboratory criteria for diagnosis can be biological (ie, a case in which brevetoxins or their metabolites are detected in clinical specimens) or environmental (ie, brevetoxins are detected in water and contaminated environmental samples by commercial laboratories using ELISA and liquid chromatography-mass spectrometry). Unfortunately, there is limited analytical capability to detect selected brevetoxins or their metabolites in urine by immunoassay. Cases are classified by diagnostic certainty: (1) suspected, if a potentially exposed person is being evaluated by health-care workers or public health officials for poisoning, but no specific credible threat is thought to exist; (2) probable, if a clinically compatible case is either thought to have had a high likelihood of brevetoxin exposure (ie, a credible threat or patient history) or has an epidemiologic link with a laboratory-confirmed case; or (3) confirmed, when laboratory tests on environmental samples are confirmatory for a clinically compatible case (17). According to the CDC, the case can also be considered “confirmed” if laboratory testing was not performed, when either sufficient clinical and nonspecific laboratory evidence of saxitoxin was present, or if the etiology of the agent is known with 100% certainty; however, this criterion is at best difficult to operationalize and seems to have been included for political reasons (ie, so the public can be reassured that the cause of an outbreak has been definitively established, even if it hasn't).
Treatment for neurologic forms of shellfish poisoning is mostly supportive and symptomatic, including monitoring of respiratory function, administration of intravenous fluids, and pain control when necessary (as in some cases of neurotoxic shellfish poisoning). Activated charcoal can be given if the patient presents within four hours of consumption, although there are no data that this alters the course of intoxication with neurologic forms of shellfish poisoning. There is no specific antidote for any of the associated toxins: domoic acid with amnesic shellfish poisoning, brevetoxin with neurotoxic shellfish poisoning, or saxitoxin with paralytic shellfish poisoning. Mannitol or brevatal (a natural antagonist of brevetoxin produced by K. brevis) may ultimately prove useful in treating neurotoxic shellfish poisoning (85).
Attempts to develop and test antitoxins to saxitoxin in experimental animals have been mixed (09). Although antitoxin combined with oxygen therapy resulted in prompt restoration of breathing and cardiovascular performance, uncompensated acidemia persisted, as did an aberrant central respiratory pattern. The benefits that were achieved were likely due to peripheral mechanisms because of the ongoing alteration of the central respiratory pattern and because the large molecular weight of the antitoxin (greater than 150,000 Da) limited entry into the CNS.
Following domoic acid intoxication in amnestic shellfish poisoning, neuropsychological assessment months or years after intoxication may show evidence of severe anterograde memory impairment with preservation of other cognitive functions (77; 15). Following severe domoic acid intoxication, electromyography and nerve conduction studies commonly show evidence of pure motor or sensorimotor neuronopathy or neuropathy (77). Electroencephalography may show diffuse slowing of background activity, periodic lateralized epileptiform discharges (PLEDs), or bitemporal independent epileptiform activity (15). Positron emission tomography may show decreased glucose metabolism in the medial temporal lobes (77; 15). In affected patients who die, neuropathological studies show neuronal necrosis and loss, predominantly in the hippocampus and amygdala, in the same pattern as that seen experimentally in animals after administration of kainic acid or domoic acid (77; 15; 67).
See also prognosis and epidemiology sections.
All contributors' financial relationships have been reviewed and mitigated to ensure that this and every other article is free from commercial bias.

Douglas J Lanska MD MS MSPH
Dr. Lanska of the University of Wisconsin School of Medicine and Public Health has no relevant financial relationships to disclose.
See ProfileNearly 3,000 illustrations, including video clips of neurologic disorders.
Every article is reviewed by our esteemed Editorial Board for accuracy and currency.
Full spectrum of neurology in 1,200 comprehensive articles.
Listen to MedLink on the go with Audio versions of each article.
MedLink, LLC
3525 Del Mar Heights Rd, Ste 304
San Diego, CA 92130-2122
Toll Free (U.S. + Canada): 800-452-2400
US Number: +1-619-640-4660
Support: service@medlink.com
Editor: editor@medlink.com
ISSN: 2831-9125
Neurobehavioral & Cognitive Disorders
Jun. 17, 2026
Peripheral Neuropathies
Jun. 11, 2026
Peripheral Neuropathies
Jun. 11, 2026
Neuro-Oncology
May. 27, 2026
Neuropharmacology & Neurotherapeutics
May. 14, 2026
General Neurology
May. 13, 2026
Neuro-Oncology
Apr. 30, 2026
Neuropharmacology & Neurotherapeutics
Apr. 23, 2026